Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Тип Mollusca. Моллюски или мягкотелые
|
Читайте также: |
Моллюски являются одной из наиболее многочисленных групп высших многоклеточных животных. В настоящее время известно около 100 тысяч современных и ископаемых видов.
Мягкое тело моллюсков не сегментировано у большинства организмов оно состоит из головы, туловища и ноги. Но голова и нога не всегда четко выделяются. На голове имеются: рот, иногда галаза и другие органы чувств. Туловище бывает двусторонне - симметричным или спирально-закрученным. Оно содержит ряд внутренних органов и защищено снаружи раковиной. Вещество для создания раковины выделяется мантией.
Начальные стадии развития моллюсков (онтогнез) напоминают развитие кольчатых червей (Annelida), вероятно малочленистые кольчатые черви и были предками моллюсков.
Подавляющее большинство моллюсков водные, преимущественно морские животные, но известны и наземные формы.
В настоящее время моллюсков делят на 2 подтипа - Amphineura (боконервные) и Cochifera (раковинные).
Представители раковинных наиболее важны для палеонтологии.
Моллюски относятся к очень древним животным- известны с начала кембрия. Они широко распространены во времени и пространстве. Изучение моллюсков играет важную роль для познания эволюции органического мира и для определения возраста пород, содержащих их остатки.
Подтип Conchigfera включает четыре класса:
1. Bivalvia (Lamellibranchiata или Pelecypoda). Двустворки (пластинчатожаберные или топороногие).
2. Scaphopoda - лопатоногие.
3. Gastropoda - брюхоногие
4. Cehpalopoda - головоногие.
Подтип Conchifera
Класс Bivalvia (Lamellibranchiata или Pelecypoda)
Тело пелеципод двусторонне - симметричное, лишенное головы, покрытое мантией и известковой двустворчатой раковиной. Нога у большинства форм развита хорошо, но у некоторых видов редуцируется.
Строение мягкого тела
Нога - удлинненое мускулистое образование, находится на брюшной стороне туловища. Нога служит для ползания, закапывания, сверления. Форма ноги килевидная, языкообразная. У некоторых пелеципод нога имеет дисковидную ползательную подошву. Паледиподы, ведущие прикрепленный образ жизни, лишены ноги( роды Mytilus, Ostrea). У палеципод имеется биссусный аппарат, который расположен либо в ноге, либо в задней части туловища. Биссусный аппарат- это щелевидная полость с довольно крупной железой, выделяющей жидкость, быстро застывающую в воде. При застывании образуются биссусные нити, которыми моллюск прикрепляется к субстрату.
Туловище располагается над ногой. В нем имеется пищеварительная система - рот, пищевод, желудок, кишечник, анус; кровеносная система, упрощенная нервная система (в виде нервных узлов), выделительная и половая системы.
Мягкое тело моллюска окружено кожистой оболочкой - мантией. У многих пелеципод складки мантии, подходя к краю раковины, плотно прилегают друг к другу, образуя мантийную полость. Полость сообщается с внешней средой только в определенных местах. Она раздвигается в области ноги и в области сифонов (длинные трубки). У некоторых моллюсков мантия открытая (род Pecten) и в полость свободно проникает вода. Мантийная полость заполнена жидкостью, близкой по составу к плазме крови. В этой полости размещены многочисленные, сложно устроенные жабры.
Палеонтологу мягкое тело пелеципод изучать обычно не приходится (если он не работает с современными формами). Основное внимание палеонтолог уделяет исследованию раковины.
Строение раковины (скелета)
Над мантией у всех пелеципод расположена раковина, состоящая из двух отдельных частей - створок. Створки соединены друг с другом на спинной стороне при помощи лигамента (связки) и замка. Створки автоматически раскрываются благодаря упругости лигамента, а закрываются (смыкаются) при помощи мускулов - замыкателей или аддукторов. Эти мускулы оставляют на внутренней стороне створок характерные мускульные отпечатки (рис. 27). Прочность и постоянство направлений смыкания обеспечивается замком.
Величина и форма раковин различные. Соотношение створок (правой и левой) непостоянно.
Раковина образуется уже на личиночной стадии. Затем первичная тонкая пластинка увеличивается в размерах и изгибается.
Размеры раковин - от долей мм почти до метра. Величина раковины и толщина створок могут служить показателем условий обитаний - крупные, массивные раковины характерны для тепловодных бассейнов. Размеры раковины меняются по мере роста. Возрастные изменения видны по линиям нарастания на поверхности створок. Форма раковины - родовой и видовой признак, она может быть шаровидной, дисковидной, уплощенной, конусовидной и конической, черенковидной или сложной, состоящей из комбинации геометрических тел.
На каждой створке различают макушку (рис. 25), обособленную начальную часть, расположенную вблизи замка.
По макушке, ориентируя ее вверх и вперед, определяют какая створка левая и какая правая.
Створки могут быть равносторонними, но у прикрепленных форм развивается неравностворчатость.
Поверхность створок бывает гладкой или скульптированной. Скульптура выражена обычно ребрами. Различают ребра радиальные (расходятся лучами от макушки) и концентрические (идут параллельно краям створок). Раковины могут иметь только радиальные или только концентрические ребра, или сочетание тех и других. Ребра бывают раздельными (двух, трех и многораздельными). У пелеципод кроме ребер наблюдаются еще и складки - концентрические или радиальные волнообразные изменения выпуклости створок (рис. 27).
Элементы внутреннего строения раковины.
След прикрепления мантии к створке называется мантийной линией. Она может быть углубленной или поверхностной, волнистой или ровной, сплошной или прерывистой, пунктирной.
У некоторых родов мантия отодвигается от края раковины, образуя изгиб или мантийный синус. Если форму его трудно описать словесно, приводят его зарисовку.
На створках под макушкой на замочной площадке у многих пластинчатожаберных имеется углубление, к которому при жизни животного прикрепляется связка или лигамент. Это хрящевидно-мускульное образование, которое выдерживает все виды напряжений - растягивающее, сжимающее, свертывающее.
Прочное смыкание створок и сохранение их положения друг относительно друга обеспечивается замочным аппаратом или замком (рис. 28). Строение замка является родовым и видовым признаком.
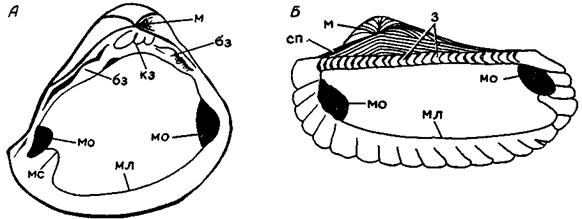
А-Mactra (правая створка), Б-Arca (левая створка): м-макушка, з-зубы, бз-боковые зубы, кз-кардинальный зуб, мо-мускульные отпечатки, мл-мантийная линия, мс-мантийный синус, сп-площадка, к которой прикреплялась связка.
Рисунок 27. Элементы внутреннего строения двустворчатых моллюсков
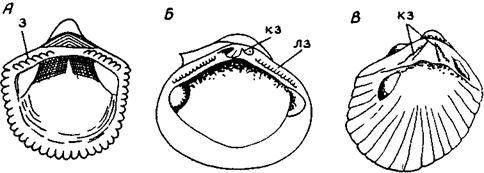
А-таксодонтный-все зубы (з) равноценны (род Pectunculus); Б-гетеродонтный (кз-кардинальный зуб, лз-латеральные зубы, род cyrena); В-схизодонтный (кардинальный зуб расцеплен, род Neotrigonia)
Рисунок 28. Некоторые типы замочного аппарата двустворок
Различают следующие виды замков двустворок.
1) Таксодонтный (рядозубый) - зубы многочисленные, чередуются с зубными ямками, расположены на обособленной замочной площадке;
2) гетеродонтный (разнозубый) - немногочисленные, различные по величине и форме зубы отделены друг от друга зубными ямками; под макушкой расположены кардинальные зубы, перпендикулярные замочному краю, по краям замочной площадки расположены латеральные или боковые зубы, параллельные замочному краю;
3) Схизодонтный (шизодонтный, расщепленнозубый) - строение аналогично гетеродонтному типу, но кардинальный зуб усложнен поперечными бороздками или насечками;
4) десмодонтный (связкозубый) - настоящие зубы отсутствуют и для поддержания связки имеются лопатовидные или ложечкообразные выступы, иногда с зубовидными бугорками;
5) дизодонтный (беззубый) - ясно выраженных зубов нет, могут присутствовать маленькие поперечные образования - первичные зубчики и первичные пластины; створки смыкаются мускулами - замыкателями и внутренней связкой.
Специфическими признаками при родовых и видовых определениях могут быть отпечатки мускулов, а также неполное смыкание створок или зияние (род Solen - зарывающаяся форма).
Историческое развитие
Двустворчатые моллюски - древние животные. Время их появления преположительно кембрий. В ордовике и силуре они были широко распространены. Кембрийские двустворки имели небольшие размеры, тонкостенные раковины, таксодонтный замок и были преимущественно морскими обитателями. В девоне появились солоноватоводные формы, а в позднем палеозое - пресноводные.
В мезозое появились новые отряды двустворок, например, Rudistae, которые участвовали в рифообразовании. Следующее существенное обновление класса произошло на рубеже мела и палеогена.
Экология и тафономия.
Двустворки - водные животные, существовавшие и существующие в морских, солоновато - и пресноводных бассейнах. Взрослая стадия жизни пластинчатожаберных связана с дном водоемов т.к. это бентосная группа. Малая подвижность обуславливает обитание двустворок на небольшом участке дна при относительном постоянстве условий жизни, поэтому ископаемые формы являются хорошими индикаторами среды.
По способу питания двутворки делятся на 4 группы: фильтраторов, грунтоедов, «хищников» и древоточцев.
В связи с придонным образом жизни двустворки ползают по дну, лежат на дне на одной из створок или прикрепляются биссусом к каким-либо предметам (камни, раковины).
Особый тип приспособления к обитанию на дне в теплых водоемах - цементация. Двустворка «прирастает» к грунту макушкой одной из створок. При большом количестве таких форм возникали рифоподобные поселения.
Отдельные двустворки могли непродолжительное время плавать (род Pecten).
Из закономерностей захоронения двустворок следует отметить:
1) в песках и галечных литорали содержится мелкораздробленный раковинный детрит;
2) по мере удаления от берега в песках двустворки захороняются в виде разрозненных створок, отсортированных по размерам и окатанности;
3) вдали от волнопрчбойной зоны встречаются целые раковины с двумя створками;
4) створки чаще всего ориентированы выпуклостью вверх и параллельно напластованию («ракушечная мостовая»).
5) в глубоководных глинистых отложениях раковины различного возраста встречаются совместно, а в их расположении нет определенной ориентировки (иногда такие тафоценозы встречаются и в мелководных отлоежениях бухт и заливов, поэтому при воссоздании палеогеографических условий следует кроме двустворок использовать и другие группы фауны).
Геологическое значение
Развитие двустворок осуществляется различными темпами: есть медленно эволюционирующие группы (pод Pteria известен с ордовика до ныне) и есть группы, которые существовали непродолжительное время (род Inoceramus - юра - мел). Для определения возраста важны роды, виды и подвиды, существовавшие краткое геологическое время и широко распространенные в пространстве. Таковы мезо-кайнозойские представители двустворок (в частности род Inoceramus). При массовых скоплениях раковин в области мелководья образуются раковинные известняки (ракушняки), которые являются прекрасным строительным и отделочным материалом.
Класс Yastropoda. Брюхоногие
Класс включает около 8000 видов современных и ~ 15 000 ископаемых животных. Брюхоногие - моллюски с асимметричным телом, обособленной головой и хорошо развитой ногой. Нога имеет широкую плоскую подошву. Тело брюхоногих - продолговатое с выпуклостью на спинной стороне. У большинства гастропод оно прикрыто раковиной - спиральной или колпочкообразной. Помещается тело в мантийной полости и представляет собой спирально- закрученный мешок, который при помощи мускулов прикрепляется к столбику внутри раковины. Стобик называется колумелла. У гастропод с колпачкообразной раковиной тело моллюска прикреплено непосредственно к внутренней стороне раковины.
Гастроподы обладают пищеварительной, кровеносной, нервной и выделительной системами. Дыхание осуществляется либо жабрами, либо легкими. Животные раздельнополые.
Строение раковины
Раковина гастропод снаружи покрыта конхиолиновым или роговым слоем, собственно раковина арагонитовая, реже кальцитовая. Внутренняя часть раковины покрыта перламутровым слоем, который иногда сохраняется в ископаемом состоянии.
Различают три типа раковин - спирально-плоскостные, спирально-конические и колпачкообразные. Наиболее разнообразны спирально-плоскостные и спирально-конические раковины. Они бывают симметричные или асимметричные (рис.30). У симметричных раковин начальные обороты находятся в конусообразных углублениях, охватываемых последующими оборотами. Обычно это пелагические, свободно плавающие формы. Асимметричные раковины образуют различной высоты завиток, состоящей из отдельных оборотов.
У многих современных брюхоногих раковина редуцирована.
Раковины гастропод имеют отверстие, через которое происходит сообщение с внешней средой.
Оно называется устьем или апертурой (рис. 29). Устье может быть разнообразным по величине и форме. Иногда по краю устья проходит сифон - трубкообразное разрастание мантии. Через сифон к жабрам подходит вода. Устье может быть закрыто крышечкой. Края устья или контур образуют околоустье или перистому.
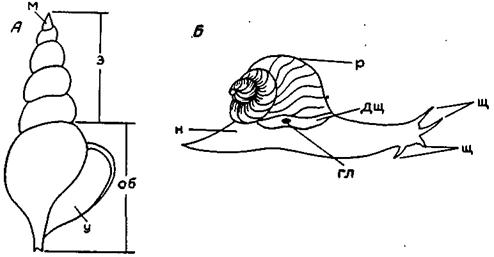
А – Tortisipha, Б – Helix pomatica: м – макушка, з – завиток, об – оборот, у – устье, р – раковина, н – нога, щ – щупальца, дщ – дыхательная щель, гл - глаз
Рисунок. 29. Основные элементы строения раковины (А) и мягкого тела (Б) асимметричных гастропод.
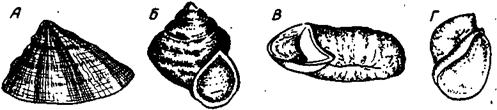
А – колпачковидная (Patella), Б – конусоспиральная – вид со стороны устья (Pleurotomaria), В – плоскоспиральная (Planorbis), Г – яйцевидная (Limnea)
Рисунок. 30. Различные формы раковин современных и ископаемых брюхоногих.
У колпачкообразных форм устье занимает все основание раковины. По характеру перистомы выделяют голостомную (с цельной перистомой) и сифоностомную(с каналом для сифона) группы гастропод.
Для взрослых особей гастропод величина раковины является постоянной при постоянных условиях обитания. При изменении внешних условий меняется и величина раковины.
Форма раковины, ее общее очертание зависит как от направления завивания (завернутый тип), так и от характера оборотов (рис. 30). Завивание может происходить как по часовой стрелке (вправо), так и против нее (влево). Обороты могут нарастать равномерно и тогда раковина имеет форму, близкую к конической. Отдельные обороты могут быть выпуклые, уплощенные, вогнутые или угловатые. По количеству оборотов различают мало - и многооборотные раковины.
Палеонтолог изучает характер оборотов последовательно - от первого к последнему, где располагается устье. Линия соприкосновения соседних оборотов называется швом.
Историческое развитие
Достоверные представители гастропод известны с кембрия, хотя некоторые исследователи полагают, что появились гастроподы в более древние эпохи. В кембрии встречаются симметричные и асимметричные раковины, а также колпачковидные. В ордовике и силуре гастроподы ведущей роли не играли. С девона и до конца палеозоя морские брюхоногие осваивали солоновато - и пресноводные водоемы, а в мезозое и кайнозое гастроподы переходили из морской среды в пресную и наземную и широко распространились.
Экология и тафономия
Большинство брюхоногих моллюсков обитает в морях с нормальной или близкой к ней соленостью Населяют они прибрежные и мелководные участки морских бассейнов. Однако отдельные виды могут существовать и на абиссальных глубинах. В личиночной стадии все брюхоногие плавают (планктон). Для взрослых особей обычен донный образ жизни.
Брюхоногие чувствительны к типу берега и характеру морского дна. На скалистом берегу, у границы прилива живут представители рода Patella. На песчаном дне живут роды, раковины которых имеют выросты и шипы, они массивны и толстостенны. На илистом берегу селятся хищные брюхоногие, которые питаются зарывающимися двустворками.
Большинство гастропод живет в сублиторельной зоне (0-70 м). В батиальной зоне (от 200 до 100 -1700 м) обитают сифоностомные формы.
Брюхоногие, дышащие легкими, могут существовать при различных температурах. Современные легочные гастроподы живут от Гренландии до Сахары и легко переносят резкие колебания температуры, благодаря способности к анабиозу.
Наземные и пресноводные брюхоногие в большинстве своем являются растительноядными организмами.
Морские гастроподы обитают в водах с соленостью от 20 до 40 - 45 %. Пресноводные формы выдерживают некоторое осолонение бассейнов - от 0,003 до 2 - 3%.
Температура существенно влияет на морфологию раковин брюхоногих. В тропических областях встречаются наиболее крупные формы, так как в морях этого пояса высока скорость осаждения CaCO3, что дает материал для строительства раковины. Понижение температуры приводит к уменьшению размеров раковины. В теплых тропических морях гастроподы разнообразны в родовом и видовом отношении, но количество особей одного вида невелико. В холодных морях отмечается обратная закономерность: количество родов и видов невелико, количество особей одного вида огромно.
Температура влияет и на размножение брюхоногих: повышение или понижение ее вызывает гибель зародыша и уже сформировавшейся личинки.
Гастроподы по типу питания подразделяются на растительноядных, плотоядных и всеядных.
Геологическое значение. Брюхоногие используются для установления возраста мезозойских и, в основном, кайнозойских отложений. Палеозойские гастроподы изучены слабо.
В связи с тем, что брюхоногие быстро реагируют на изменение внешней среды изменением морфологии раковины, их используют для палеогеографических реконструкций.
Классификация брюхоногих моллюсков
Основана на строении органов дыхания, нервной системы и ноги. Выделяют три подкласса: переднежаберные (Prosobranchia), заднежаберные (Opistobranchia) и легочные (Pulmonata).
Подкласс Prosobranchia.
К подклассу относятся наиболее древние брюхоногие. Они имеют нервный перекрест и одну или две жабры, расположенные впереди сердца. Раковина улитковидная, колпачковидная или дискоидальная. Подкласс делится на три отряда:
1) Archaeogastropoda (археогастроподы);
2) Mesogastropoda (мезогастроподы);
3) Neogastropoda (неогастроподы).
Подкласс Opistobranchia
Не имеет перекреста нервных тяжей, единственная жабра позади сердца или ее нет и дыхание осуществляется всей поверхностью тела. Раковина плохо развита, иногда она охватывается мантией и становится внутренней или исчезает. Распространение - карбон - ныне.
Подкласс представлен отрядом Pteropoda (крылоногие). Это брюхоногие, ведущие планктонный образ жизни. Нога преобразуется в две крыловидные лопасти. Раковина тонкостенная или отсутствует. Распространение - палеоген - ныне.
Подкласс Pulmonata.
Дыхание легочной полостью Раковина тонкостенная, спирально-плоскостная или спирально-коническая, иногда отсутствует. Обитают на суше или в пресноводных бассейнах. Многочисленная группа. Распространение - карбон - ныне.
Класс Cephalopoda. Головоногие.
К классу относятся наиболее высокоорганизованные, крупные, очень подвижные, свободно плавающие или ползающие хищные моллюски, распространенные в океанах и морях с нормальной соленостью.
Из современных цефалопод известны кальмары, каракатица, осьминоги, аргонавты и наутилус («жемчужный кораблик»).
Тело головоногих имеет двустороннюю симметрию и состоит из головы и туловища (рис. 31). На голове расположен рот, окруженный щупальцами, и сложные глаза. Тело покрыто мантией, которая на брюшной стороне образует мантийную полость. Нервная система высокоразвитая. Хорошо развиты пищеварительная и кровеносная системы. Жабры расположены в мантийной полости. Нога видоизменена. Передняя ее часть образует щупальца, задняя - воронку. Воронка служит для выведения продуктов обмена из мантийной полости, а также для передвижения. Животное двигается задним концом вперед.
Раковина у большинства головоногих представляет собой трубку, открытую с одного конца. Состоит раковина из жилой камеры и фрагмакона (рис. 31). В жилой камере помещается тело моллюска. Фрагмакон подразделен перегородками на многочисленные камеры (гидростатические). Они заполнены газом и водой, от заднего конца тела отходит кожистая трубка - сифон, протягивающийся через все камеры (рис. 31), он регулирует содержание газа и жидкости в камерах.
Раковина головоногих моллюсков прямая или согнутая, плоскоспиральная или спирально-коническая. У многих современных форм раковина сильно редуцирована или отсутствует.
Головоногие появились в кембрии, широко распространены в палеозое и мезозое. Многие имеют важное стратиграфическое значение.
В зависимости от характера раковины головоногие подразделяются на два подкласса - Ectocoсhlia (наружнораковинные) и Endocochlia (внутрираковинные).
Подкласс Ectocochlia Наружнораковинные.
Раковина имеет разнообразную форму, поделена поперечными перегородками на камеры. Тело моллюска помещается в последней камере остальные заполнены смесью газов. Все камеры сообщаются друг с другом через сифон (рис. 31).
Все Ectocochlia - обитатели морей с нормальной соленостью.
Подкласс делится на несколько надотрядов, из которых наибольшее значение имеют Nautiloidea и Ammonoidea.
Надотряд Nautiloidea. Наутилоидеи.
Наутилоидеи - головоногие с наружной раковиной, коротким мешковидным телом, большим количеством щупалец и воронкой (рис. 31). Устье от щелевидного до широкого округлого. Перегородки между камерами прямые или изогнутые. Сифон находится в центре раковины, имеет двухслойную оболочку. У некоторых моллюсков имеются дополнительные специальные отложения внутри камер и внутри сифона. Распространение -кембрий - ныне.
Строение и состав раковины
Раковина наутилоидей состоит из наружного органического слоя (периостракума), фарфоровидного слоя и внутреннего перламутрового. В ископаемом состоянии обычно сохраняется фарфоровидный (кальцитовый) слой, органический и перламутровый слои сохраняются редко.
У ископаемых наутилоидей преобладают формы с прямой раковиной. Реже встречаются роговидные, яйцевидные и спирально-свернутые. Спирально-свернутые подразделяются на планоспиральные (свернуты в одной плоскости) и турбоспиральные (свернуты по принципу винта).
Планоспиральные раковины бывают двух типов: необъемлющие или эволютные и объемлющие или инволютные. У эволютных раковин каждый последующий оборот не закрывает предыдущий, у инволютных раковин каждый последующий оборот закрывает не только предыдущий, но и все более ранние обороты. Существуют раковины промежуточного типа, которые на ранних стадиях развиваются как эволютыне, а на более поздних - как инволютные.
Скульптура раковин представлена линиями нарастания (тонкие поперечные струйки) и ребрами.
Камеры раковины наутилоидей отличаются друг от друга только по размерам, а жилая камера еще и расширяется к устью. Стенки воздушных камер утолщены. Дополнительные отложения находятся на вентральной или брюшной стороне.
Перегородки, отделяющие камеры друг от друга обычно направлены в сторону устья. Линия соединения края перегородки с внутренней поверхностью раковины называется перегородочной (линией). Часто она бывает сложно изогнутой или лопастной.
Сифон - задний конец тела наутилоидей оттянут в виде шнуровидного или конического образования, окруженного оболочкой и сохраняющегося в ископаемом состоянии. Это образование и называется сифон (рис. 31). Сифон проходит через отверстия всех перегородок, соединяя воздушные камеры. Внутри сифона могут быть дополнительные отложения.
Историческое развитие
Наутилоидеи известны из нижнекембрийских отложений (роды Volbortella, Salterella). В среднем кембрии известен род Vologdinella. Некоторые палеонтологи считают, что указанные моллюски не относятся к наутилоидеям. Типичные наутилоидеи определены из позднекембрийских отложений Сибири, Китая, Северной Америки. Эти наутилоидеи очень мелкие, со сжатым поперечным сечением и широким сифоном. В ордовике наблюдался расцвет наутилоидей. В этот период появились все известные отряды и семейства, увеличились размеры раковин, более разнообразными стали их форма, скульптура и строение сифона. В девоне, карбоне и перми количество наутилоидей несколько сокращается. В мезозое и кайнозое наутилоидеи представлены только одним отрядом. Среди современных наутилоидей известен только род Nautilus (рис. 31).
Экология и тафономия.
В настоящее время наутилоидеи обитают только в тропической зоне западной части Тихого океана (о-ва Фиджи, Новая Гвинея, новые Гебриды, Новая Каледония, Филиппины), но их пустые раковины разносятся течениями и встречаются у берегов Японии, Индии, Австралии и даже Мадагаскара. Наутилусы живут на глубинах от нескольких метров до 500 - 700 м. По способу питания они относятся к хищникам и ведут придонный образ жизни, хотя и являются хорошими пловцами мезозойские и кайнозойские формы вели аналогичный образ жизни.
Образ жизни палеозойских наутилоидей трудно реконструировать. Палеонтологи предполагают, что прямые наутилоидеи жили в мелкой воде, активно плавали, находясь в горизонтальном положении. Некоторые формы были пассивно плавающими (планктон) или ползающими по дну. В захоронениях наутилоидеи встречаются совместно с аммоноидеями. Обычно это единичные экземпляры, скопления раковин не обнаружены.
Биологическое и геологическое значение
Наутилоидеи единственная группа головоногих, существующих с кембрия (?) поныне. Все остальные головоногие, очевидно произошли от наутилоидей. Велико значение наутилоидей для определения геологического возраста палеозойских отложений, особенно ордовика и силура, частично девона.
Каменноугольные и пермские наутилоидеи редко встречаются в массовом количестве.
В мезозое важное значение имели меловые наутилоидеи. Более поздние формы мало изучены и не используются для определения возраста.
| <== предыдущая страница | | | следующая страница ==> |
| ТИП ARCHAEOCYATHI. АРХЕОЦИАТЫ | | | Надотряд Ammonoidea. Аммоноидеи |
Дата добавления: 2014-04-28; просмотров: 1315; Нарушение авторских прав

Мы поможем в написании ваших работ!