Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Вирусы как мутагены
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РФ
Филиал ЧОУ ВПО «БИЭПП» в г. Мурманске
ЮРИДИЧЕСКИЙ ФАКУЛЬТЕТ
РЕФЕРАТ
Вирусы как мутагены.
Дисциплина: Концепция современного естествознания
Группа: ЭЗ – 31C - 11
Студент: Давыдова Анна Николаевна
Преподаватель: Ф.И.О. преподавателя полностью
(ученая степень, звание)
Дата регистрации работы_________
Оценка: _______
Домашний адрес:
г. Ковдор,
ул. Горняков,
д. 4, кв. 28
Ковдор
СОДЕРЖАНИЕ:
1. Введение
2. Основные термины и понятия
3. Факторы способствующие мутагенезу
4. Механизм вирусных мутаций
5. Вирусы-мутагены
6. Роль мутагенов в эволюции
7. Список использованных источников
ВВЕДЕНИЕ
Основы понимания роли мутаций в эволюции были заложены в 20-х гг. 20 в. Работами советского генетика С. С. Четверикова, английских учёных Дж. Холдейна и Р. Фишера и американского учёного С. Райта, положивших начало развитию эволюционной генетики. Было показано, что все наследственные изменения, служащие материалом для эволюции, обязаны мутациям (комбинативная изменчивость, возникающая путём образования новых сочетаний генов при скрещивании, в конечном счёте, тоже есть следствие мутаций, обусловливающих генетические различия скрещивающихся особей).
Способность к мутациям присуща всем живым существам. Мутагены универсальны, т.е. могут приводить к мутациям у других организмов. В отличие от модификаций, мутации ненаправленный: один и тот же мутагенный фактор, действующий с одинаковой интенсивностью на организмы с идентичным генотипом (например, однояйцевых близнецов), может вызывать у них разные мутационные изменения. Вместе с тем различные мутагенные факторы могут вызывать у далеких в генетическом отношении организмов подобные наследственные изменения. Степень выражения мутаций в фенотипе не зависит от интенсивности и длительности действия мутагенного фактора: за кратковременной слабого действия мутагенного фактора мутационные изменения могут быть выражены более, чем за длительной и сильной, но в последнем случае возрастает частота мутаций.
Мутации являются одним из источников наследственной изменчивости – важного фактора эволюции организмов. Большинство из них вредна для живых существ, поскольку снижает приспособленность к условиям существования, приводит к разнообразным нарушениям процессов жизнедеятельности и наследственных заболеваний. Но нейтральные мутации, которые почти не выливают на жизнеспособность организмов, при определенных изменений окружающей среды могут оказаться полезными.
ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ
Вирусы (лат. Virus — «яд») — неклеточный инфекционный агент, который может воспроизводиться только внутри живых клеток.
Мутация (лат. Mutatio — изменение) — стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) преобразование генотипа, происходящее под влиянием внешней или внутренней среды.
Мутагены (от «мутация» и др.-греч. Γεννάω — рождаю) — химические и физические факторы, вызывающие наследственные изменения — мутации.
Мутагенами могут быть различные факторы, вызывающие изменения в структуре генов, структуре и количестве хромосом. По происхождению мутагены классифицируют на эндогенные, образующиеся в процессе жизнедеятельности организма и экзогенные — все прочие факторы, в том числе и условия окружающей среды.
Мутагенез — внесение изменений в нуклеотидную последовательность ДНК (мутаций).
Инсерционный мутагенез (лат. Inserto — вставлять, вкладывать; лат. Mutatio — изменение и греч. Genesis — происхождение, развитие) — мутационное изменение генома вследствие вставок нуклеотидных последовательностей мобильных генетических элементов или вирусов, а также с помощью трансфекции или трансгеноза; в результате И.м. может происходить или частичная, или полная инактивация генов.
Вирус-индуцируемый инсерционный мутагенез (лат. Virus — яд и лат. Inductio — возбуждение; лат. Inserto — вставлять, вкладывать; лат. Mutatio — изменение и genesis — происхождение) — мутагенез в клетках, зараженных вирусом, который способен внедрять свой геном в различные участки генома клетки-хозяина, вызывая этим инсерционные мутации (напр., так возникла мутантная окраска шерсти у мышей, зараженных вирусом лейкоза Молони).
Точечная мутация — тип мутации в ДНК или РНК, для которого характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен, инсерции или делеции одного или нескольких нуклеотидов.
ФАКТОРЫ, СПОСОБСТВУЮЩИЕ БИОЛОГИЧЕСКОМУ МУТАГЕНЕЗУ
В конце 60-ых и в начале 70-ых годов сразу несколько групп ученых обнаружили, что вирусы являются фактором мутагенеза . Дальнейшие исследования позволили установить, что практически все известные вирусы были способны индуцировать цитогенетические изменения в клетках человека и экспериментальных животных. Такие данные были получены относительно вируса кори, гриппа, паротита, краснухи, оспы, полиомиелита и многих других.
Особое беспокойство ученых вызвало то, что не только инфекционные формы вируса, но и живые вакцины также способны индуцировать повышение числа клеток с нарушениями в числе и структуре хромосом. Кроме того, исследование на дрозофилах, проведенное известным киевским ученым С.М. Гершензоном и его учениками показало, что вирусы способны вызывать генные мутации. Причем, даже не патогенные для дрозофилы вирусы человека обладали ярко выраженным мутагенным эффектом. Ученый предположил, что мутагенным действием обладает нуклеиновая кислота вируса. Особое внимание привлекли данные, свидетельствующие о специфическом поражении определенных локусов (участков) хромосом. Так, например, вирус кори вызывал нарушения преимущественно во 2-ой хромосоме, а вирус гриппа в 6 и 9. В основном нарушения локализовались в зонах так называемой повышенной ломкости хромосом и местах сосредоточения онкогенов, неверное функционирование которых может приводить к развитию раковых опухолей. Анализ уровня цитогенетически измененных клеток и состояния иммунореактивности организма у больных пациентов (грипп, корь, клещевой энцефалит, полиомиелит), позволил установить, что имеется четко выраженная зависимость – чем больше цитогенетически измененных Т-лимфоцитов, тем выраженнее Т-иммунодепрессия у данного пациента. В связи с этим было высказано предположение, что иммуноцит с нарушениями в хромосомном аппарате не может выполнять “возложенные” на него функции.
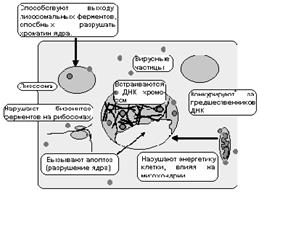
Рис. 1 Внутриклеточные факторы инсерционного мутагенеза
В случае инфекционного процесса при размножении возбудителя внутри клетки, как правило, наблюдается конкуренция между инфектом и клеткой за предшественники нуклеиновых кислот. Поскольку инфект размножается очень быстро, то клетка-хозяин может недополучать “строительный материал”. При вступлении в стадию синтеза ДНК инфицированная клетка испытывает дефицит структурных компонентов ДНК, и это также может обусловить появление ошибок при биосинтезе наследственных молекул.
Известно, что в ликвидации инфекта принимают активное участие лейкоциты – макрофаги, нейтрофилы, Т- и В – лимфоциты. Эти клетки стремятся уничтожить проникший паразитарный агент или инфицированную им клетку, при этом образуются самые разнообразные “активные” молекулы – перекиси, синглентный кислород а также некоторые ферменты, которые могут способствовать возникновению поломок в генетическом аппарате клеток инфицированного организма. Внимание привлекают антинуклеарные антитела, которые образуются при многих аутоиммунных конфликтах, а также при инфекционных заболеваниях. Показано, что они способны проникать и соединяться с хроматином ядра. Некоторые из них специфичны, например, при склеродермии образуются антитела против кинетохоров хромосом, что может обусловить неверное расхождение хромосом при делении клеток.
МЕХАНИЗМ ВИРУСНЫХ МУТАЦИЙ
Обычно ДНК точно копируется при процессе репликации и сохраняется неизменной между двумя последовательными репликациями. Но изредка происходят ошибки и последовательность ДНК меняется – эти ошибки называются мутациями.Мутация это устойчивое наследуемое изменение ДНК, независимо от его функциональной значимости. Это определение подразумевает изменение в первичной нуклеотидной последовательности, а изменения иного рода, например метилирование , обычно относят к эпигенетическим событиям .
Мутации в соматических клетках, возможно, вызывают процессы старения , рак и другие, менее существенные изменения в организме.
Мутации в половых клетках родителей наследуются детьми.
Представление об устойчивости мутаций в целом остается верным, но открытие динамических мутаций, обусловленных увеличением числа тринуклеотидных повторов, показывает, что некоторые мутации изменяются при делении соматических или зародышевых клеток. Одни мутации летальны, и они не могут передаваться следующему поколению, а другие не столь опасны и сохраняются в потомстве.
С точки зрения эволюции мутации обеспечивают достаточное генетическое многообразие, чтобы позволить видам приспособиться к условиям окружающей среды путем естественного отбора.
Каждый генетический локус характеризуется определенным уровнем изменчивости, т. Е. присутствием различных аллелей, или вариантов последовательностей ДНК, у разных индивидуумов. Применительно к гену, аллели разделяются на две группы – нормальные, или аллели дикого типа, при которых функция гена не нарушена, и мутантные, приводящие к нарушению работы гена. В любых популяциях и для любых генов аллели дикого типа являются преобладающими. Под мутацией понимают все изменения в последовательности ДНК, независимо от их локализации и влияния на жизнеспособность особи. Таким образом, понятие мутации является более широким по сравнению с понятием мутантного аллеля .
В научной литературе часто встречающиеся в популяциях варианты последовательностей генов, не приводящие к заметным нарушениям функций, обычно рассматриваются как нейтральные мутации или полиморфизмы , тогда как понятия «мутация» и «мутантный аллель» зачастую употребляются как синонимы.
Мутации могут захватывать участки ДНК разной длинны. Это может быть единственный нуклеотид, тогда мы будем говорить о точковой мутации, или же протяженный участок молекулы. Кроме того, учитывая характер изменений, мы можем говорить о заменах нуклеотидов , делециях и вставках (инсерциях) и о инверсиях. Процесс возникновения мутаций называют мутагенезом . В зависимости от факторов, вызывающих мутации, их разделяют на спонтанные и индуцированные . Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды. Спонтанные мутации в эукариотических клетках возникают с частотой 10-9-10-12 на нуклеотид за клеточную генерацию.
Индуцированными называют мутации, возникающие в результате мутагенных воздействий в экспериментальных условиях или при неблагоприятных воздействиях окружающей среды. Среди важнейших мутагенных факторов, прежде всего, необходимо отметить химические мутагены – органические и неорганические вещества, вызывающие мутации, а также ионизирующее излучение .
Между спонтанными и индуцированными мутациями нет существенных различий, Большинство спонтанных мутаций возникает в результате мутагенного воздействия, которое не регистрируется экспериментатором.
Классификация мутаций основана на молекулярных процессах их возникновения.
Геномы большинства ДНК-содержащих вирусов (за исключением поксвирусов ) транскрибируются в ядре клетки-хозяина с помощью клеточной РНК-полимеразы II . Синтез и процессинг мРНК у этих вирусов практически полностью осуществляется за счет клеточных механизмов (несколько вирусных белков участвуют только в регуляции транскрипции).
У многих ДНК-содержащих вирусов транскрипция происходит в три этапа, во время которых избирательно считываются сверхранние, ранние и поздние гены.
Нуклеокапсиды герпесвирусов проникают в цитоплазму вместе с белками матрикса, затем этот комплекс транспортируется вдоль микротрубочек к порам ядерной оболочки и далее в ядро. Белки матрикса вместе с клеточными факторами транскрипции запускают транскрипцию сверхранних генов. У герпесвирусов продукты сверхранних генов, так называемые сверхранние белки, играют роль трансактиваторов и необходимы для начала транскрипции ранних генов. Однако у других ДНК-содержащих вирусов транскрипция ранних генов может происходить и без участия сверхранних белков.
Большинство ранних генов кодируют белки, осуществляющие репликацию вирусной ДНК и запуск транскрипции поздних генов. Поздние гены кодируют главным образом структурные белки, необходимые для сборки вирусов и их выхода из зараженной клетки. Транскрипция поздних генов зависит от репликации ДНК и прекращается в присутствии ингибиторов репликации.
Для каждого семейства ДНК-содержащих вирусов характерен уникальный механизм репликации ДНК.
Геном герпесвирусов представлен линейной ДНК, которая в зараженной клетке замыкается в кольцо. Кольцевые молекулы вирусной ДНК реплицируются по механизму «катящегося кольца» с образованием длинных линейных конкатемерных молекул, состоящих из многих копий генома. Затем конкатемерные молекулы расщепляются на полноценные геномы. В репродукции герпесвирусов участвуют вирусные ДНК-полимераза и ферменты, увеличивающие внутриклеточную концентрацию дезоксинуклеозидтрифосфатов – структурных элементов ДНК.
Репликация линейных геномов аденовирусов происходит при участии вирусной ДНК-полимеразы и нуклеотид-белкового комплекса, выполняющего роль праймера .
Двухцепочечные кольцевые геномы паповавирусов реплицируются с сохранением кольцевой структуры. Репликация осуществляется клеточными ферментами, а для запуска репликации необходим ранний вирусный белок – Т-антиген . Изредка геномы паповавирусов (например, папилломавирусов ) встраиваются в хромосому зараженной клетки, что приводит к усиленной экспрессии вирусных белков и неконтролируемому делению клетки. Интеграция паповавирусов в клеточный геном может стать причиной злокачественного новообразования , например рака шейки матки .
В отличие от репродукции других ДНК-содержащих вирусов, репродукция поксвирусов полностью происходит в цитоплазме. Эти вирусы содержат факторы транскрипции, РНК-полимеразу, а также набор ферментов, обеспечивающих процессинг мРНК (присоединение 7-метилгуанозина, полиаденилирование). Геном поксвирусов имеет уникальное строение: комплементарные цепи двухцепочечной ДНК на обоих концах ковалентно соединены между собой, так что в действительности геном состоит из одноцепочечной кольцевой молекулы ДНК. Кроме того, в геноме имеются инвертированные концевые повторы. Репликация вирусной ДНК начинается с расщепления одной из цепей в области инвертированных концевых повторов; затем одновременно на обоих концах генома начинается синтез комплементарной цепи, который осуществляет вирусная ДНК-полимераза. Праймером служит прилегающий к месту расщепления короткий участок расщепленной цепи (самозатраванный механизм). Поксвирусы, как и герпесвирусы , кодируют несколько ферментов, способствующих репликации ДНК путем увеличения концентрации дезоксирибонуклеозидтрифосфатов.
ВИРУСЫ-МУТАГЕНЫ
· РНК-содержащие вирусы — вирусы, у которых генетический материал содержится в одно- или двухцепочечной линейной РНК, а не в ДНК. У позвоночных существует около 30 семейств или отдельных родов РНК-с.в.: Retroviridae, Reoviridae, Picornaviridae, Coronaviridae и др. Размеры РНК-геномов РНК-с.в. варьируют от 7х103 кДа у пикорнавирусов до 2х105 кДа у парамиксовирусов. Большинство РНК-с.в. размножаются путем образования копий РНК без участия промежуточных ДНК-матриц, и поэтому их репликация может происходить в клетках с ингибированным синтезом ДНК; эти вирусы кодируют собственную РНК-репликазу. Среди них различают РНК-с.в. с положительным РНК-геномом (вирусы полиомиелита и клещевого энцефалита у животных, вирус табачной мозаики у растений) и отрицательным РНК-геномом (вирусы гриппа, кори, бешенства, желтой карликовости картофеля и др.); у первых геном представлен мРНК, у вторых — анти-мРНК. РНК-с.в. семейства Retroviridae размножаются с помощью обратной транскрипции РНК ревертазой с образованием комплементарной ДНК. Отдельные семейства РНК-с.в. вызывают неопластические заболевания, другие семейства являются причиной таких патологий человека и животных, как воспалительные заболевания верхних дыхательных путей, гастроэнтерит, менингит и др. Некоторые РНК-с.в. животных используются в качестве векторов для генной терапии. Существуют также РНК-с.в. растений (вирус бороздчатости ствола яблони, вирус кольцевой пятнистости табака), грибов (вирус L-A, Cryphonectria hypovirus 1-EP713) и бактерий (фаги MS2, Q-B). Впервые инфекционность вирусных РНК было продемонстрирована Х. Френкель-Конратом с соавтором в 1957 г.
· ДНК-содержащие вирусы (лат. Virus — яд) — вирусы, генетический материал которых представлен одно- или двухцепочечной молекулой ДНК (линейной или кольцевой формы) размером от 2 до 560 тыс. п.н. Геном может быть представлен одной молекулой ДНК (несегментированный) или несколькими молекулами ДНК (сегментированный). ДНК-с.в. образуют 24 семейства и 3 неклассифицированных рода вирусов с двуцепочечной ДНК и 5 семейств и 2 неклассифицированных рода вирусов с одноцепочечной ДНК. У позвоночных насчитывается 11 семейств ДНК-с.в. (иридовирусы, поксвирусы, асфавирусы, герпесвирусы, аденовирусы, полиомавирусы, папиломавирусы, гепаднавирусы, парвовирусы, цирковирусы и анелловирусы). Самыми простыми из перечисленных выше вирусов являются парвовирусы, цирковирусы и анелловирусы; их геном представлен одноцепочечной ДНК длиной 2—6 тыс. н. Самые крупные — покс-вирусы, размер их генома достигает 375 тыс. п.н. Почти у всех ДНК-с.в. ДНК синтезируется в ядре зараженной клетки, там же созревают их вирионы; лишь у поксвирусов все стадии размножения происходят в цитоплазме. Многие ДНК-с.в. вызывают разнообразные заболевания человека и животных. Различные аденовирусы часто используются в качестве векторов при генной терапии. К ДНК-с.в. относятся также бакуловирусы (см. Бакуловирусы), размножающиеся в клетках членистоногих, которые широко используются в биотехнологии. Имеются ДНК-с.в. и у растений (напр., вирус полосатости кукурузы, мозаичный вирус сои).
РОЛЬ МУТАГЕНОВ В ЭВОЛЮЦИИ
Основы понимания роли мутагенов в эволюции были заложены в 20-х гг. 20 в. работами советского генетика С. С. Четверикова, учёных Дж. Холдейна и P. Фишера и американского учёного С. Райта, положивших начало развитию эволюционной генетики. Было показано, что все наследств, изменения, служащие материалом для эволюции, обязаны мутации (комбинативная изменчивость, возникающая путём образования новых сочетаний генов при скрещивании, в конечном счёте, тоже есть следствие мутации, обусловливающих генетические различия скрещивающихся особей). В отличие от модификаций, мутации не являются однозначной реакцией на вызывающее их воздействие; один и тот же мутагенный фактор приводит к возникновению разнообразных мутаций, затрагивающих те или иные признаки организма и изменяющих их в разных направлениях. Поэтому сами по себе мутации не имеют адаптивного характера. Однако постоянно возникающие у любого вида живых существ мутации, многие из которых к тому же длительно сохраняются в популяции в скрытом виде (рецессивные мутации), служат резервом наследств, изменчивости, который позволяет естественному отбору перестраивать наследств. признаки вида, приспосабливая его к меняющимся условиям среды (изменению климата или биоценоза, переселению в новый ареал и т. п.). Адаптивность эволюционных изменений - следствие сохранения естественным отбором носителей тех мутаций и их сочетаний, которые оказываются полезными в данной обстановке. При этом мутации, бывшие в одних условиях вредными или нейтральными, могут оказаться полезными в изменившихся условиях. Наибольшее значение для эволюции имеют генные мутации. Несмотря на относит, редкость мутаций каждого гена, общая частота спонтанных генных мутаций весьма значительна, т. к. генотип многоклеточных организмов состоит из десятков тысяч генов. В результате ту или иную генную мутацию несёт большая доля образуемых организмом гамет или спор (у высших растений и животных эта доля достигает 5-30% ), что создаёт предпосылки для эффективного действия естественного отбора. Хромосомные перестройки, затрудняющие рекомбинацию,- инверсии и транслокации - способствуют репродуктивной изоляции отдельных групп организмов и их последующей дивергенции; дупликации ведут к увеличению числа генов в генотипе и возрастанию их разнообразия вследствие происходящей затем дифференциации генов в дуплицированных участках хромосом. Полиплоидия играет большую роль в эволюции растений; при этом, помимо репродуктивной изоляции, она в ряде случаев восстанавливает плодовитость бесплодных межвидовых гибридов.
С разработкой способов искусств, мутагенеза открылась возможность значит, ускорения селекции - селекционерам стал доступен гораздо больший исходный материал, чем при использовании редких спонтанных мутаций. В 1930 сов. учёные А. А. Сапегин и Л. H. Делоне впервые применили ионизирующую радиацию в селекции пшеницы. В дальнейшем методами радиационной селекции были выведены новые высокоурожайные сорта пшеницы, ячменя, риса, люпина и других сельско-хозяйственных растений, ценные штаммы микроорганизмов, используемых в промышленности. В селекции с хорошими результатами применяются и химические мутагены.
Геномные мутации, хромосомные перестройки и генные мутации - причина многих наследственных заболеваний и врождённых уродств у человека. Поэтому ограждение человека от действия мутагенов - важнейшая задача. Огромное значение в этом отношении имело осуществлённое по инициативе СССР запрещение испытаний ядерного оружия в атмосфере, загрязняющих окружающую среду радиоактивными веществами. Очень важно тщательное соблюдение мер защиты человека от радиации в атомной индустрии, при использовании радиоактивных изотопов, рентгеновских лучей и т. п. Необходимо изучение возможного мутагенного действия различных новых лекарственных средств, пестицидов, химических препаратов, применяемых в промышленности, и запрещение производства тех из них, которые окажутся мутагенными. Профилактика вирусных инфекций имеет значение и для защиты потомства от мутагенного действия вирусов.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. wikipedia.org
2. allbest.ru
3. humbio.ru
4. podelise.ru
5. mysteryvirus.ru
6. bse.slovaronline.com
7. med-books.info
8. www.vse-pro-geny.com
| <== предыдущая страница | | | следующая страница ==> |
| Вирус иммунодефицита человека | | | ВИРУСОЛОГИЯ |
Дата добавления: 2015-06-30; просмотров: 1242; Нарушение авторских прав

Мы поможем в написании ваших работ!