Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Численность N пред
|
Читайте также: |
B d
Рис. 3.6. Возрастная пирамида популяции полевки
Рис. 3.5. Типы возрастных пирамид
Рис. 3.4. Разные типы кривых выживания
Рис. 3.3. Диапазон изменений плотностей популяций у разных видов млекопитающих
Генетические характеристики. Генотип - совокупность всех генов данного организма.Еще в начале прошлого века популяцию рассматривали как совокупность генетически неоднородных особей, противопоставляя ее генетически чистым линиям. В дальнейшем этот подход приобрел генетико-эволюционный смысл и в современном представлении популяция рассматривается как элементарная единица эволюционного процесса. С этих позиций главный критерий популяции - способность к свободному обмену генетической информацией.
Характеристики популяционной группы
Популяция характеризуется рядом признаков, причем единственным носителем этих признаков является группа, а не особи. Вот некоторые из таких свойств популяции: численность, рождаемость, смертность, возрастная структура, распределение в пространстве. Популяции обладают также генетическими и фенотипическими характеристиками, непосредственно связанными с условиями обитания. Каждая особь в популяции отличается одна от другой, но все они вместе представляют сложный биологический комплекс особей, распределенных по своим индивидуальным качествам по кривой, близкой к нормальному распределению.
Основные характеристики популяции можно разделить на две самостоятельные группировки:
· характеристики, определяющие предпосылки для самостоятельного существования и развития популяции;
· характеристики, определяющие биологическую специфичность (морфологическую и физиологическую) слагающих популяцию особей.
К генетическим характеристикам в первую очередь относят: адаптивность, репродуктивную (дарвиновскую) приспособленность и непрерывность (т.е. вероятность оставления потомков на протяжении длительного периода времени).
Изменения внешних условий
 | |||
 | |||
ГЕНОТИП (ФЕНОТИП)
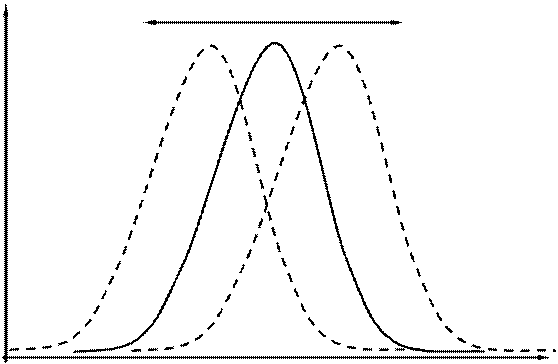
Рис. 3.2. Распределение особей различных генотипов (фенотипов) в популяции.
Популяция как элементарная природная совокупность особей является "взаимоскрещивающимся единством" и состоит из особей разных генотипов, распределенных по кривой близкой к нормальному распределению (рис. 3.2). Большинство природных популяций распадается на микропопуляции - более мелкие группы особей, могущие иметь различные частоты встречаемости генотипов. Они могут быть временно изолированны (в отдельные сезоны года), но вследствие миграций и расселения молодняка и других процессов связаны между собой и имеют единый генофонд. В большинстве случаев между соседними популяциями осуществляется постоянный обмен особями, а значит и генетической информацией. Этим определяется их генетическая близость, но при этом различия по морфологическим и физиологическим особенностям между ними сохраняются.
Генетическое единство - основное свойство популяции, определяющее все ее особенности и пути ее развития.
Фенотипические характеристики. Фенотип – это совокупность различных морфологических и физиологических особенностей растений и животных, которая формируется под влиянием конкретных условий существования.
К фенотипическим характеристикам популяции в первую очередь относятся скорость роста и развития, общие размеры тела (вес), плодовитость, пропорции тела, окраска и т.д. Даже в близко расположенных популяциях, но обитающих в неодинаковых условиях, могут достаточно ярко проявляться морфологические и физиологические различия.
Если популяции обитают в среде с постоянным однонаправленным действием каких - либо факторов среды, живые организмы, составляющие ее, приобретают специфические фенотипические и генетические особенности. Именно эти особенности и определяют в последующем направления естественного отбора и во многом - эволюцию вида. При этом происходит смещение максимума частоты встречаемости фенотипа, детерминированного генетически в ту или иную сторону (рис. 3.2). Одним из основных параметров популяции, прежде всего, является ее численность или плотность.
Плотность. Плотность популяции - среднее число особей на данной территории, отнесенное к некоторой единице пространства или объема. Например: 150 растений сосны на 1 га, или 0,5 г циклопов в 1 м3 воды характеризуют плотность популяции этих видов.
| Хищные | Рысь Пума | Плотоядные |
| Лиса Медведь | Всеядные | |
| Растительноядные | Бурундук Белка | Поедающие семена |
| Полевки Лесной сурок Олень | Поедающие зеленую массу |
0.001 0.01 0.1 1.0 10 100
Биомасса, кг/га
Она никогда не бывает постоянной и определяется соотношением рождаемости и смертности, а также процессов эмиграции и иммиграции. В процессе размножения происходит рост популяции, смертность же приводит к сокращению численности. Экологическая плотность - число особей (или биомасса) на единицу заселенного пространства (т.е. доступной площади или объема, которые реально могут быть заняты популяцией).
Численность особей в каждой из популяций варьирует в зависимости от размеров особей того или иного вида и ареала популяции. У насекомых, мелких травянистых растений она может достигать сотни тысяч и более. Популяции же крупных животных и больших древесных растений бывают небольшими по численности (рис. 3.3).
Численность популяции непостоянна. В природе не отмечено ни одной популяции, которая бы оставалась стабильной даже на протяжении короткого отрезка времени. Основными причинами ее изменения являются изменения условий существования. Это связано с воздействием абиотических факторов на интенсивность роста и размножения, на смертность, на скорость развития. Например, известно, что динамика популяций белки, зайца, многих видов птиц зависит в большинстве случаев от урожая кормов. В настоящее время изменения численности популяции нередко зависят от деятельности человека. Немалую роль в этом процессе играет уничтожение хищниками жертв, конкурентные взаимоотношения, влияние паразитов на жизнедеятельность хозяев и т. д.
Однако, несмотря наколебания, численность популяции изменяется не беспредельно. Популяция, как и любая биологическая система, обладает способностью к саморегуляции. Для популяции каждого вида всегда имеются верхние и нижние пределы плотности, за границы которых она выходить не может. При благоприятном сочетании всех условий плотность популяции удерживается на каком-то оптимальном уровне, незначительно отклоняясь от него в ту или другую сторону. Такие колебания численности носят регулярный характер и четко отражают реакцию популяции на конкретные условия среды. Падение плотности ниже оптимальной приводит к ослаблению защитных реакций популяции, снижает ее плодовитость, могут привести к вымиранию конкретной популяции и вызывать ряд других отрицательных явлений. Повышение плотности сверх оптимальной также неблагоприятно сказывается на популяции, поскольку при этом иссякает кормовая база, сокращается жизненное пространство, появляются вспышки различных заболеваний и т. д.
Различают непериодические, редко наблюдаемые и периодические, постоянные, колебания численности естественных популяций. К непериодическим колебаниям численности, а соответственно и плотности популяции можно отнести вспышки массового размножения среди насекомых (непарный шелкопряд в южной части России 1879 г.), грызунов и т. д. Резкий подъем численности наблюдается у популяций, оказавшихся в новом местообитании. Классическими примерами этому может служить массовое размножение кроликов в Австралии, колорадского жука в Европе.
Периодические изменения численности популяций совершаются обычно в течение нескольких лет или одного сезона. Циклические изменения численности с подъемами в среднем через четыре года зарегистрированы у таких животных как песец, полярная сова, у леммингов. Как правило, циклические изменения численности полярной совы коррелируют с периодическими изменениями численности леммингов - основной пищи сов. Численность сов, возрастающая по мере увеличения количества пищи, затем также быстро снижается по мере ее уменьшения. При этом численность популяция сов резко уменьшается в результате миграции. Эти колебания численности так регулярны, что орнитологи США могут точно предсказать нашествие полярных сов каждые три или четыре года. Поскольку эти птицы хорошо заметны и появляются повсюду вокруг городов, они привлекают к себе большое внимание. Хорошо известные примеры циклических изменений численности у жуков-короедов (Питермен, 1978). Благодаря деятельности жуков, которые изреживают загущенный древостой, малопригодный для лесозаготовок и обитания в нем животных, создаются более ценные для человека леса. Деятельность этих жуков можно рассматривать как одно из средств ведения хозяйства, а не как вредителей леса. В изолированных лесах, по-видимому, следовало бы допускать вспышки численности этих жуков.
Сезонные колебания численности характерны для многих видов насекомых, грызунов, птиц, а также для мелких водных организмов.
Размножение. Динамика численности и плотности популяции находятся в тесной зависимости от размножения и смертности, а так же от способности слагающих их особей совершать миграции.
Рождаемость (плодовитость) - характеризует процесс появления на свет новых особей любого организма независимо от того, рождаются ли они, вылупляются из яиц, прорастают из семян или появляются в результате деления, что приводит к увеличению численности популяции. Этот термин используется также и при анализе популяций человека (в демографии). Обычно рождаемость выражают либо как скорость, определяемую путем деления общего числа вновь появившихся особей на время (общая рождаемость), либо как число вновь появившихся особей в единицу времени на 1 особь в популяции (удельная рождаемость).
В живых организмах заложена огромная потенциальная возможность к размножению. Подсчитано, что бактерии делятся каждые 20 минут. При таком темпе одна клетка за 36 часов дает потомство, которое может покрыть сплошным слоем всю планету. Один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут. В действительности же такая громадная плодовитость организмов никогда не реализуется. В связи с этим различают максимальную или потенциальную рождаемость и экологическую или реализованную рождаемость.
Потенциальная рождаемость - это теоретически возможная максимальная скорость образования новых особей в идеальных условиях, когда отсутствует действие лимитирующих экологических факторов, а размножение сдерживается только физиологическими возможностями организма.
Экологическая или реализованная рождаемость означает увеличение численности популяции при фактических условиях среды. Эта величина не постоянна и изменяется в зависимости от размерного и возрастного состава популяции и физических условий среды.
Например, в колонии чайки отложено 510 яиц (потенциальная рождаемость) из них вылупилось 265 птенцов, реализованная плодовитость составила 52% от потенциальной. Мучные хрущики отложили 1200 яиц – вылупилось 713 особей, что составило всего 60%. У капустной белянки в результате гибели яиц, гусениц и куколок по разным причинам (болезни, паразиты, хищники и др.) на свет появляется лишь 0,32 % взрослых бабочек от числа отложенных яиц.
Рождаемость каждого вида определяется как приспособление, обеспечивающее пополнение популяций, а значит, ее жизнестойкость. Поэтому естественно, что у менее приспособленных к неблагоприятным условиям видов высокая смертность в молодом возрасте компенсируется значительной плодовитостью. В зависимости от взаимоотношений между родителями и потомками существует две противоположные стратегии размножения. Виды животных и растений, у которых забота о потомстве выражена явно - смертность их потомства на ранних стадиях развития низка, а сами они имеют низкую плодовитость. Потомство видов, у которых отсутствует забота о детенышах, имеют высокую смертность, а их родители - высокий репродуктивный потенциал (как компенсацию за высокую смертность потомков). Для видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая потенциальная и низкая экологическая рождаемость. В благоприятных условиях рождаемость обычно низкая а в неблагоприятных – высокая. Этим объясняется более высокая рождаемость животных в зонах умеренного климата по сравнению с тропическими видами.
Смертность. Численность и плотность популяции зависят также от ее смертности. Смертность популяции - это количество особей, погибших за определенный период. Она, как и рождаемость, изменяется в зависимости от условий среды, возраста и состояния популяции и выражается в процентах к начальной или чаще к средней ее величине. Выделяют: экологическую смертность - гибель особей, наблюдаемую в конкретных условиях среды (эта величина непостоянна и изменяется в зависимости от условий среды и состояния самой популяции); теоретическую минимальную смертность - величина, постоянная для популяции; она представляет собой гибель особей в идеальных условиях, при которых популяция не подвергается действию лимитирующих факторов. Даже в самых лучших условиях особи будут умирать от старости. Этот возраст определяется физиологической продолжительностью жизни, которая часто превышает среднюю экологическую продолжительность жизни.
Различают четыре основных типа возрастных изменений смертности (рис. 3.4).
I. Первый тип смертности характеризуется повышенной гибелью взрослых, главным образом старых особей. Наблюдается он у многих видов насекомых, личинки которых обитают в почве, воде, древесине или других местах с благоприятными условиями.
II. Ступенчатая кривая (выживание резко меняется при переходе от одной стадии жизненного цикла к другой).
 |
III. К третьему относится животные и растения, смертность которых одинакова во всех возрастах. Она описывается уравнением прямой. Такая смертность встречается очень редко и только у популяций, постоянно находящихся в оптимальных условиях.
IV. Четвертый тип смертности отличается повышенной гибелью особей на ранних стадиях развития и свойствен большинству растений и животных. Максимальная гибель многих растений происходит в стадии прорастания семян и всходов, а животных - в личиночной фазе или в молодом возрасте (у многих рыб до взрослой фазы доживает 1- 2 % от числа выметанной икры, у насекомых - 0,3-0,5 % от отложенных яиц).
Чаще всего процессы смертности оцениваются по кривым выживания. Они выражают зависимость числа выживших из 100 или 1000 особей от их возраста.
Форма кривой выживания связана со степенью заботы о потомстве и другими способами защиты молоди. Так, кривые выживания пчел и дроздов (которые заботятся о потомстве) значительно менее вогнуты, чем у кузнечиков и сардин (которые не заботятся о потомстве). У этих видов это обстоятельство компенсируется значительно большим числом откладываемых яиц.
Форма кривой выживания очень часто изменяется так же в зависимости от плотности популяции. На мало заселенной территории больше шансов выжить. То же самое можно сказать и относительно человеческих популяций, где высокая плотность не всегда бывает благоприятна для отдельного индивидуума. Многие экологи считают, что быстрый рост и высокая плотность в популяциях человека опасны не столько для выживания, сколько для качества жизни индивидуума. И это несмотря на то, что экологическая продолжительность жизни человека значительно увеличилась благодаря достижению современной медицины, улучшению питания и т. д. Однако физиологическая продолжительность жизни человека, по-видимому, не увеличилась, поскольку в наше время до 100 лет доживает не больше людей, чем в минувшие столетия.
На динамику численности популяции большое влияние оказывают миграции отдельных особей, т. е. обмен индивидуумов популяций. Это происходит при переселении их из одного местообитания в другое. В основном миграции совершаются при подрастании молодняка и его расселении. В результате миграций устраняется избыток особей в одной популяции и компенсируется недостаток в другой.
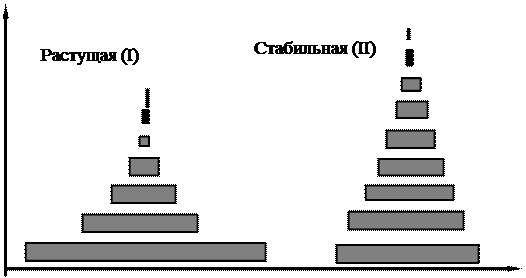
Возрастная структура популяции. Возрастная структура популяции является ее важной характеристикой, которая влияет как на рождаемость, так и на смертность. Соотношение разных возрастных групп в популяции определяет ее способность к размножению в данный момент и показывает, что можно ожидать в будущем. У большинства видов репродуктивная способность их членов изменяется с возрастом. В начальный период (который может продолжаться всего несколько минут, как у микроорганизмов, или несколько лет, как у человека и многих видов деревьев) организмы размножаться не способны. За этим периодом следует период постепенного повышения способности к размножению, достигающей максимума, а затем начинается спад, доходящий, в конечном счете, до полного прекращения. Соответственно в популяции обычно выделяют три экологические возрастные группы (фазы): 1). предрепродуктивную; 2). репродуктивную; 3). пострепродуктивную (рис. 3.5). Длительность этих возрастных фаз по отношению к общей продолжительности жизни варьирует у разных организмов. У современного человека три этих возраста приблизительно одинаковы, на каждый из них приходится около трети жизни (рис.3.7). У первобытных людей пострепродуктивный период был намного короче. Для многих животных и растений характерен очень длительный предрепродуктивный период. В большинстве природных популяций особи гибнут, не достигнув пострепродуктивного периода. Однако люди, домашние животные, а также животные, которых содержат в зоопарках, живут в более благоприятных условиях, а поэтому длительность пострепродуктивного периода может быть значительной. Обычно в быстро растущих популяциях значительную долю составляют молодые особи (рис. 3.5 (I), рис. 3.6 (I)) в популяции, находящейся в стационарном состоянии (в стабильных популяциях), возрастное распределение более равномерно (рис. 3.5 (II), рис. 3.6 (II)), а в популяции, численность которой снижается, будет содержаться большая доля старых особей (рис. 3.5, (III)). Возрастная структура популяции может изменяться и без изменения ее численности. Любая популяция, находящаяся в стабильной среде при благоприятных условиях, в конце концов, приближается к некоторому стабильному возрастному распределению.
 |  | | |||






 I II III
I II III
 | |||||||
 | |||||||
 | |||||||
 | |||||||


|
Временные отклонения от него могут быть вызваны лишь необычными увеличениями рождаемости или смертности, после чего происходит возврат к стабильной ситуации.
 | |||||
 | |||||
| |||||
 |
Рис. 3.7. Возрастная структура популяции человека, рассчитанная отдельно для мужчин и женщин в Швеции и Коста – Рике. Население Швеции увеличивается медленно, поэтому основания пирамиды узкое. Процент старших возрастных классов (40 - 60 лет) велик. Быстрый рост населения Коста – Рики сформировал пирамиду с широким основанием и высоким процентом населения моложе 25 лет (Ricklefs, 1979).
Изучение половозрастного состава популяции имеет большое значение. Один из основных факторов, влияющих на размеры популяции, - это процент особей, погибающих до достижения половой зрелости (смертность в дорепродуктивном периоде). В пределах вида эта величина гораздо более изменчива, чем рождаемость. У промысловых птиц, и пушных зверей отношение сеголеток к старшим возрастным группам служит показателем тенденции изменения численности популяции. Это отношение определяется во время охотничьего сезона (осенью или зимой) по выборке из популяции, полученной охотниками или промысловиками. В общем, высокая численность молоди по сравнению с численностью взрослых указывает на то, что сезон размножения сложился успешно; и если смертность молоди не будет чрезмерной, то можно ожидать увеличения популяции в следующем году.
Рост популяции. Такие показатели, как рождаемость, смертность и возрастное распределение, весьма важны, но каждый их них сам по себе мало что дает; в частности, по каждому их них нельзя судить о росте популяции как целого, о характере изменений, которые произойдут при изменении условий среды, о возможностях популяции.
Внутренняя скорость роста.Если среда не налагает никаких ограничений (пространство, пища и другие организмы не оказывают лимитирующего действия), то скорость роста популяции для данных микроклиматических условий становится максимальной и постоянной. Значение скорости роста в этих условиях характеризуется соответствующей возрастной структурой и представляет собой единственный показатель наследственно обусловленной способности к росту - r.
dN/dt = rN, (1)
где: dN/dt- скорость изменения численности (скорость роста); r - скорость изменения численности в расчете на одну особь в единицу времени; N– численность.
Кривая роста с учетом действия лимитирующих факторов, как правило, имеет S - образный вид. Проинтегрировав уравнение (1) получаем экспоненциальную зависимость
Nt= N0 ert, (2)
где N0- численность в начальный момент времени, Nt - численность в момент времени t и e - основание натурального логарифма. После логарифмирования:
r = dN/(Ndt), (3)
r=(ln Nt – ln N 0)/t, (4)
если вместо Nt и N0 подставить Nt1 и Nt2 , вместо t - (t1 - t2) в фазе нелимитирующего роста получим скорость роста между двумя промежутками времени.

 b - const
b - const
 |
Удельная рождаемость (b)
 |
 Удельная смертность (d)
Удельная смертность (d)
 | |||
 |

 b d
b d
| <== предыдущая страница | | | следующая страница ==> |
| | |
Дата добавления: 2014-03-21; просмотров: 820; Нарушение авторских прав

Мы поможем в написании ваших работ!