Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
График ВПСП
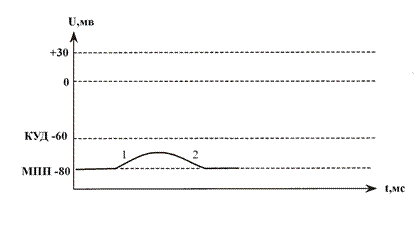
1) медленная деполяризация, 2) медленная реполяризация.
ВПСП является местным, нераспространяющимся возбуждением, обладающим свойствами ЛО. Вследствие генерации ВПСП появляется относительная разность потенциалов между частично деполяризованной постсинаптической мембраной и наиболее возбудимым поляризованным участком центрального нейрона - аксонным холмиком (начальным сегментом аксона). Поэтому, между возбужденным и невозбужденными участками мембраны нейрона начинают циркулировать местные ионные токи, которые являются непосредственной причиной разрядной деятельности в области аксонного холмика.
Чем больше частота афферентных ПД, тем больше в синапсе выделится медиатора. Поэтому выше будет амплитуда ВПСП на постсинаптической мембране, а значит больше частота ПД в области аксонного холмика.
Центральные химические синапсы, как и мионевральные, характеризуются четырьмя основными свойствами:
1) односторонним поведением возбуждения,
2) синаптической задержкой,
3) низкой лабильностью,
4) высокой утомляемостью.
Высокая утомляемость химических синапсов обусловлена тремя основными причинами:
1) уменьшением запасов медиатора в нервном окончании,
2) снижением чувствительности постсинаптической мембраны к медиатору,
3) нарушением ресинтеза медиатора в синапсе.
Синаптическая задержка передачи сигналов и низкая лабильность обусловлены затратами времени:
1) на выделение медиатора из нервного окончания,
2) на диффузию медиатора через синаптическую щель,
3) на взаимодействие медиатора со специфическими рецепторами постсинаптической мембране.
Эфферентное звено рефлекторной дуги морфологически представлено аксонами нейронов, которые формируют центробежные нервные волокна. Его основная функция - передача от нервного центра к эффектору эфферентных ПД, в которых закодирована команда к действию.
Эффектор - это исполнительный рабочий орган, деятельность которого управляется нервным центром. Так, исполнительными органами могут быть мышцы, сосуды, железы организма.
Общая схема рефлекторной дуги
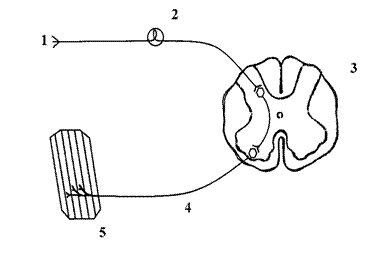
1) рецептор, 2) афферентное звено, 3) нервный центр, 4) эфферентное звено, 5) эффектор.
Для формирования рефлекторной реакции эффектора с момента раздражения рецепторов требуется определенное время. Интервал времени от начала действия раздражителя на рецепторы до появления ответной рефлекторной реакции эффекторов, называется общим временем рефлекса. Это время требуется для возбуждения рецепторов, проведения возбуждения по афференту, нервному центру, эфференту и для возбуждения исполнительного органа. Чем больше сила раздражителя, тем меньше общее время рефлекса.
Время, в течение которого возбуждение проводится по нервному центру, называют центральным временем рефлекса. Центральное время рефлекса зависит от количества центральных синапсов в рефлекторной дуге. В полисинаптической рефлекторной дуге центральное время рефлекса больше, чем в моносинаптической.
Деятельность эффекторов направлена на достижение полезного для организма приспособительного результата (ППР), который характеризуется специфическими сомато-вегетативно-эндокринными параметрами. Информация о совершенном действии и параметрах ППР по каналам обратной афферентации снова поступает в нервный центр.
Обратная афферентация морфологически представлена сенсорными нейронами аксоны которых формируют афферентные нервные волокна. Она является тем дополнительным и необходимым звеном, которое обеспечивает замыкание рефлекторной дуги и превращение ее в рефлекторное кольцо. Основная функция обратной афферентации - передача информации о совершении действия и о параметрах достигнутого ППР в нервный центр. Благодаря этому происходит коррекция его управляющей деятельности.
Схема рефлекторного кольца
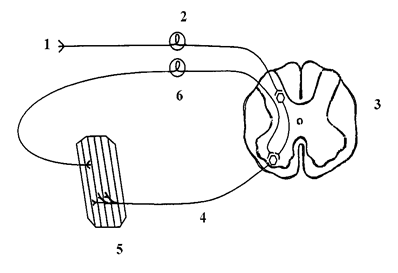
1) рецептор, 2) афферентное звено, 3) нервный центр, 4) эфферентное звено, 5) эффектор, 6) обратная афферентация.
Рефлексы отличаются большим многообразием и подразделяются на различные группы по ряду признаков.
В зависимости от расположения рецепторов выделяют экстерорецептивные и интерорецептивные рефлексы. Экстерорецептивные рефлексы вызываются раздражением рецепторов внешней поверхности тела. Интерорецептивные рефлексы могут быть висцерорецептивными и проприорецептивными. Висцерорецептивные возникают при раздражении рецепторов внутренних органов. Проприорецептивные рефлексы обусловлены раздражением рецепторов скелетных мышц, суставов, связок и сухожилий.
По характеру ответной реакции различают моторные, секреторные и сосудодвигательные рефлексы. В моторных рефлексах исполнительным органом являются мышцы. Их разновидностью являются сосудодвигательныерефлексы, которые обеспечивают изменение просвета сосудов. Секреторныерефлексы регулируют деятельность желез.
В зависимости от локализации нервных центров различают 6 основных видов рефлексов:
1) спинальные, в которых участвуют нейроны спинного мозга,
2) бульбарные, осуществляемые при обязательном участии нейронов продолговатого мозга,
3) мезэнцефальные, осуществляемые при участии нейронов среднего мозга,
4) мозжечковые, в которых принимают участие нейроны мозжечка,
5) диэнцефальные, в которых участвуют нейроны промежуточного мозга,
6) кортикальные, в осуществлении которых принимают участие нейроны коры больших полушарий.
По количеству центральных синапсов в рефлекторной дуге рефлексы подразделяются на моносинаптические и полисинаптические. Рефлекторные дуги моносинаптических рефлексов имеют два нейрона - афферентный чувствительный и эфферентный, между которыми расположен один центральный синапс. Рефлекторные дуги полисинаптических рефлексов имеют, как минимум, три нейрона: афферентный, вставочный и эфферентный.
В зависимости от продолжительности ответной реакции рефлексы могут быть:
1) фазические - быстрые и короткие,
2) тонические - продолжительные и медленные.
По биологической значимости для организма рефлексы могут быть:
1) пищевые, обеспечивающие пополнение запасов питательных веществ,
2) половые, направленные на продолжение рода,
3) оборонительные, обеспечивающие защиту организма,
4) ориентировочные, которые проявляются реакцией на новый раздражитель (рефлекс «что такое?»),
5) локомоторные, обеспечивающие движение тела.
По биологической направленности выделяют три вида рефлексов:
1) рефлексы, направленные на уравновешивание организма с внешней средой,
2) рефлексы, направленные на уравновешивание организма с внутренней средой,
3) рефлексы, направленные на продолжение рода.
И.П. Павлов выделил три основных принципа организации рефлекторных реакций организма:
1) последовательный детерминизм,
2) структурности и функции,
3) анализ и синтез.
Согласно принципу последовательного детерминизма (причинности) возбуждение по рефлекторной дуге распространяется последовательно - от рецепторов к эффекторам. При этом активация каждого последующего звена рефлекторной дуги обусловлена возбуждением предыдущего.
В соответствии с принципом структурности функции каждый морфологический элемент рефлекторной дуги выполняет специфическую функцию: рецепторы – восприятие раздражителя, афферентные нервные волокна – проведение возбуждения в ЦНС, нервный центр – анализ и синтез сигналов, эфферентные нервные волокна – проведение возбуждения к исполнительному органу.
Сущность анализа заключается в разделении поступающей в ЦНС информации на простые сенсорные сигналы. Синтез сводится к интеграции сенсорных сигналов и формированию команды для исполнительных органов. Это происходит на основе выбранной в процессе анализа наиболее важной (приоритетной) информации.
Являясь основным механизмом деятельности ЦНС, рефлексы обеспечивают поддержание гомеостаза и быстрое приспособление организма к постоянно изменяющимся условиям среды обитания. Это достигается путем сложной интеграции биоэлектрических процессов во всех отделах ЦНС.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Основные вопросы: Свойства нервных центров: одностороннее и замедленное проведение возбуждения, повышенная утомляемость, низкая лабильность, способность к иррадиации и суммации, последействие, трансформация ритма, пластичность, повышенная чувствительность к недостатку кислорода и питательных веществ.
Интеграция нервных процессов и рефлекторная деятельность ЦНС, лежащие в основе приспособительных реакций организма, во многом определяется общими свойствами нервных центров:
1) односторонним проведением возбуждения,
2) замедленным проведением возбуждения,
3) низкой лабильностью,
4) повышенной утомляемостью,
5) способностью к иррадиации,
6) способностью к суммации,
7) последействием (пролонгированием),
8) трансформацией ритма,
9) высокой пластичностью,
10) способность к тонической активности,
11) повышенной чувствительностью к недостатку питательных веществ и кислорода.
Одностороннее проведение возбуждения – это способность нервных центров проводить возбуждение только в одном направлении - от афферентов к эфферентам.
Если раздражать электрическим током афферент, то в эфферентных нервных волокнах возникает серия ПД. Однако, при раздражении эфферента, в афферентных волокнах возбуждение не возникает. Одностороннее проведение сигналов обусловлено возможностью передачи возбуждения в центральных химических синапсах только от пресинаптической мембраны к постсинаптической.
Схема эксперимента, доказывающего одностороннее проведение возбуждения в нервном центре
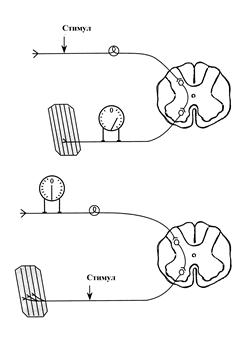
Замедленное проведение возбуждения - это свойство нервных центров, передавать возбуждение с низкой скоростью.
Между моментом поступления нервного импульса к окончанию аксона и формированием ВПСП на постсинаптической мембране проходит около 0,5 мс. Время от поступления нервного импульса к окончанию аксона до формирования ВПСП называется синаптической задержкой.
Низкая лабильность - это свойство нервных центров, характеризующееся относительно небольшой частотой импульсации, которую они способны воспроизвести без искажения ритма стимуляции афферента.
Лабильность нервных центров не превышает 50-70 ПД/с. Это обусловлено синаптической задержкой передачи возбуждения в центральных химических синапсах.
Повышенная утомляемость - это свойство нервных центров, которое проявляется в постепенном снижении или полном прекращении рефлекторного ответа при продолжительном раздражении афферентных волокон.
Снижение работоспособности нервных центров связано с нарушением передачи возбуждения в межнейронных химических синапсах и обусловлено повышенной утомляемостью центральных химических синапсов.
Иррадиация – это свойство нервного центра передавать возбуждение в ЦНС на другие нервные центры при достаточно сильном воздействии раздражителя на рефлексогенную зону.
Благодаря распространению возбуждения на несколько нервных центров рефлекторная реакция приобретает генерализованный характер. Иррадиации препятствуют процессы торможения, которые в нормальных условиях ограничивают распространение возбуждения в ЦНС.
В основе иррадиации возбуждения в ЦНС лежит морфологический принцип дивергенции (дихотомического ответвления аксонов нейронов), который заключается в способности клеток нервного центра устанавливать многочисленные синаптические контакты с нейронами других нервных центров
Схема иррадиации возбуждения в ЦНС
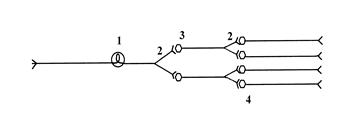
1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентные нейроны.
Способность к суммации - это свойство нервных центров формировать рефлекторную реакцию при действии на рецепторы раздражителей, которые являются подпороговыми для нервного центра.
Различают два вида суммации: последовательную (временную) и одновременную (пространственную). Последовательная суммация наблюдается при взаимодействии возбуждений, которые поступают в нервный центр с коротким интервалом времени друг за другом по одним и тем же афферентам.
Если на рецептор действует один пороговый для рецептора и афферента, но подпороговый для возбуждения нервного центра раздражитель, то разрядная деятельность в области аксонного холмика центрального нейрона, а значит и рефлекторный ответ, не возникают. Однако, при последовательном действии серии таких раздражителей, в центральном синапсе увеличивается выделение медиатора. Вследствие этого на постсинаптической мембране происходит суммация ВПСП и начинается циркуляция местных ионных токов, сила которых достаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и для возникновения ответной рефлекторной реакции эффектора.
Пространственнаясуммация возникает при взаимодействии возбуждений, одновременно приходящих в нервный центр по нескольким афферентам. При одновременном действии раздражителя на несколько рецепторов одного рецептивного поля, сигналы от которых конвергирует к одному нейрону, увеличивается выделение медиатора в центральных синапсах. Вследствие этого происходит суммация ВПСП на постсинаптической мембране, и начинают циркулировать местные ионные токи, сила которых достаточна для генерации серии эфферентных ПД в области аксонного холмика, что приводит к рефлекторному ответу исполнительного органа.
Последействие - это способность нервного центра продлевать рефлекторный ответ после прекращения раздражения рецепторов.
Известны два основных механизма последействия: кратковременный и долговременный. Кратковременное рефлекторное последействие связано со следовой деполяризацией постсинаптической мембраны нейрона после прекращения раздражения афферента. После прекращения раздражения рецепторов на постсинаптической мембране центрального синапса сохраняется следовая деполяризация в виде ВПСП, который поддерживает высокую возбудимость эфферентного нейрона. В это время за счет теплового движения молекул в синаптическую щель может самопроизвольно выделиться нескольких квантов медиатора, что приводит к образованию миниатюрного ВПСП, который суммируется со следовым потенциалом. В результате этого продолжается циркуляция местных ионных токов и поддерживается разрядная деятельность эфферентного нейрона, а следовательно и рефлекторная реакция.
За счет кратковременного механизма разрядная деятельность нейрона после прекращения раздражения афферента продлевается на несколько десятков мс. По мере уменьшения амплитуды ВПСП импульсная активность нейрона постепенно уменьшается, а затем прекращается совсем.
Долговременноерефлекторноепоследействие обусловлено реверберацией - циркуляцией возбуждения по замкнутым нейронным цепям в нервных центрах.
После прекращения раздражения возбуждение продолжает циркулировать по замкнутым нейронным цепям–«ловушкам», возвращаясь к одному и тому же эфферентному нейрону. Это обеспечивает поддержание на высоком уровне его возбудимости и продолжение разрядной деятельности. Благодаря нейронным «ловушкам» возбуждение может длительно циркулировать в нервном центре до тех пор, пока не наступит утомления одного из синапсов или же активность нейронов не будет приостановлена процессом торможения.
Схема организации нейронной ловушки
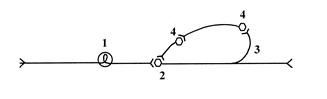
1) афферент, 2) эфферентный нейрон, 3) коллатераль, 4) вставочные нейроны.
Трансформация ритма - это свойство нервных центров изменять частоту проходящих через них импульсов.
Частота импульсации при прохождении через нервный центр может увеличиваться или уменьшаться. Способность нервных центров увеличивать частоту проходящих через них импульсов, называют мультипликацией ритма. Выделяют два основных механизма мультипликации: кратковременный и долговременный.
Кратковременнаямультипликация, как и последействие, связана с суммацией после одиночного стимула на постсинаптической мембране миниатюрного и следового ВПСП.
Долговременнаямультипликация обусловлена распространением возбуждения по дихотомически разветвляющимся коллатералям аксонов афферентов и вставочных нейронов, которые затем конвергируют (сходятся) к одному эфферентному нейрону. Вследствие схождения к эфференту нескольких вставочных нейронов, которые связаны с одним афферентом, нервный центр на одиночное раздражение афферента отвечает серией эфферентных ПД.
Схема нейронной организации долговременной мультипликации
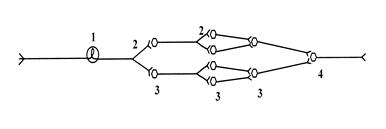
1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентный нейрон.
Пластичность - это способность нервных центров изменять свое функциональное назначение в зависимости от особенностей условий деятельности.
Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками. Она является рабочим механизмом обучения. Наибольшей пластичностью обладает кора больших полушарий головного мозга. При повреждении какой-либо зоны коры через некоторое время другие ее участки могут брать на себя выполнение функции утраченного отдела.
Способность к тонической активности заключается в постоянной минимальной импульсной активности нервных центров, которая поддерживается за счет афферентных сигналов от рефлексогенных зон. Тоническая активность нервных центров обеспечивает тонус большинства исполнительных органов и их постоянную готовность к деятельности даже в состоянии относительного покоя.
Повышенная чувствительность к недостатку питательных веществ и кислорода обусловлена высоким уровнем метаболизма нервной ткани. Специфичность обмена веществ в нейронах состоит в исключительности использования в качестве питательного материала глюкозы, тогда как другие клетки используют также различные моносахариды, жирные кислоты и аминокислоты. Кроме того, в нейронах преобладает окислительное фосфорилирование, требующее непрерывной доставки О2.
При относительно небольшой массе, составляющей не более 2% от веса тела, мозг человека потребляет до 20% всего поступившего в организм кислорода и 17% глюкозы. Интенсивность потребления глюкозы настолько велика, что в нейронах не успевают образоваться запасы гликогена. Поэтому уменьшение доставки глюкозы к ЦНС быстро влечет за собой нарушение функции нервных центров.
Интенсивность потребления нервными клетками кислорода составляет около 50 мл в мин. Потребляя такое большое количество О2, мозг и особенно кора больших полушарий высокочувствительны к его недостатку. Поэтому уже через 5-7 секунд после прекращения кровообращения в мозге человек теряет сознание, а через 5-6 минут нервные клетки больших полушарий подвергаются необратимым биологическим изменениям.
Центры ствола мозга менее чувствительны к недостатку кислорода: их функция может восстановиться после 20 минут полного прекращения кровообращения. Центры спинного мозга еще более выносливы. Их функция восстанавливается даже через 30 минут после полного прекращения притока крови.
При гипотермии - понижении температуры тела, вследствие снижения уровня обмена веществ, ЦНС дольше переносит недостаток О2 и питательных веществ. Гипотермия при температуре тела 25-280С повышает устойчивость коры больших полушарий к кислородной недостаточности с 5 до 15-20 минут.
| <== предыдущая страница | | | следующая страница ==> |
| РЕФЛЕКТОРНЫЙ ПРИНЦИП ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ (ЦНС) | | | Рыночная среда создает основу для организации предпринимательской деятельности |
Дата добавления: 2014-08-09; просмотров: 966; Нарушение авторских прав

Мы поможем в написании ваших работ!