Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Генетическая рекомбинация без гомологии: процессы, ведущие к перестройкам в геноме
Генетически рекомбинации без гомологии либо использует очень ограниченную гомологию между рекомбинирующими ДНК, либо вообще обходится без нее. Они внесли существенный вклад в эволюцию генетического материала, но наиболее ярко их биологическое значение проявляется в онтогенетических перестройках геномов, играющих важную роль в жизнедеятельности вирусов, бактерий и эукариот. Эти рекомбинационные системы - сайт-специфическая рекомбинация, транспозиции и незаконная рекомбинация - в корне отличаются от гомологичной рекомбинации по механизмам и наборам контролирующих их генов [109, Глазер В.М., Соросовский образовательный журнал, 1998].
САЙТ-СПЕЦИФИЧЕСКАЯ РЕКОМБИНАЦИЯ
Сайт-специфическая рекомбинация происходит между специфическими последовательностями ДНК в пределах очень коротких участков гомологии, обычно 15-30 п.н. Она широко распространена у прокариот и низших эукариот. Сайт-специфическая рекомбинация обеспечивает:
· интеграцию (включение) ДНК умеренных фагов в хромосомы бактерий,
· инверсию отдельных участков ДНК в хромосомах бактерий и бактериофагов и в так называемой 2-микронной плазмиде дрожжей.
· Редкий, если не единственный, но зато жизненно важный пример сайт-специфической рекомбинации у многоклеточных животных - перестройки в последовательностях ДНК, кодирующих иммуноглобулины.
Важно отметить, что все изученные ферменты, непосредственно осуществляющие сайт-специфическую рекомбинацию, являются сайт-специфическими топоизомеразами I. По уровню гомологии их аминокислотных последовательностей и механизму катализируемых реакций их разделяют на две группы:
· представителями первой группы являются уже рассмотренная нами интеграза фага лямбда, интегразы других умеренных фагов и белок, катализирующий сайт-специфическую рекомбинацию в плазмиде дрожжей,
· ко второйотносят инвертазы бактерий и фагов и некоторые другие ферменты сайт-специфической рекомбинации.
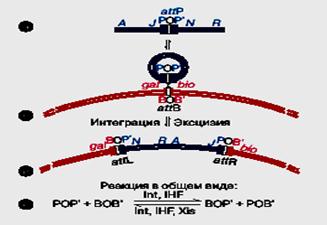 Рисунок 62. Схема сайт-специфической рекомбинации у фага лямбда (Глазер В.М. 1998).
Рисунок 62. Схема сайт-специфической рекомбинации у фага лямбда (Глазер В.М. 1998).
|
Рассмотрим некоторые из самых известных примеров сайт-специфической рекомбинации. Наиболее изучена она у умеренного бактериофага лямбда, осуществляемые топоизомеразами I группы:
1. После инфекции в клетку E. coli линейная вирионная двуцепочечная ДНК фага (рис. 62, а) замыкается в кольцо (рис. 62, б) за счет имеющихся на ее концах комплементарных одноцепочечных последовательностей.
2. Последующее развитие фага может идти по пути интеграции в хромосому бактерии между генами gal и bio (рис. 62, в). Интеграция происходит путем рекомбинации между особыми att (attachment)-сайтами: attP в хромосоме фага и attB в хромосоме бактерии. Интегрированный фаг называется профагом. Он фланкирован рекомбинантными сайтами attL (левый) и attR (правый).
3. Вырезание (эксцизия) профага из хромосомы происходит в обратной последовательности событий. Запись реакций в обобщенном виде представлена на рис. 62, г. Из нее видно, что в интегративной рекомбинации участвуют сайты attP и attB, продукт фагового гена int (интеграза) и белок IHF (Integration Host Factor) E. coli. Для эксцизии необходимы сайты attL и attR, те же белки и еще продукт фагового гена xis. Сайты attP и attB неравноценны. Первый устроен сложно. При размере около 270 п.н. он состоит из центральной части О и двух примыкающих к ней частей: P и P'. Внутри сайта attP располагаются участки связывания с интегразой и белками IHF и Xis. Сайт attB устроен проще. Его размер всего 23 п.н., гомология с attP ограничена центральной частью из 15 п.н., в которой имеются два сайта связывания с интегразой. Интеграза является топоизомеразой типа I, но только сайт-специфической. В результате образования комплекса интегразы с attP-сайтом и белком IHF формируется сложно уложенная нуклеопротеидная структура, которая захватывает сайт attB. В этой структуре последовательности ДНК att-сайтов связываются за счет взаимодействий между субъединицами интегразы и затем подвергаются согласованному расщеплению.
Как и все топоизомеразы I, интеграза делает разрыв в одной цепи каждого дуплекса, и в месте разрыва образуются 3'-P и 5'-OH-концы ДНК. Фермент ковалентно связывается с 3'-P-концом, благодаря чему энергия разорванной фосфодиэфирной связи сохраняется, и для последующего замыкания разрыва, осуществляемого тем же ферментом, не требуется дополнительной энергии. При этом интеграза может осуществлять рекомбинацию между att-сайтами путем замыкания фосфодиэфирной связи с 5'-OH-концом из другого дуплекса, то есть путем обменов 5'-OH-концами.
Такая рекомбинация между дуплексами, не требующая дополнительной энергии, называется консервативной. Молекулярный процесс интегративной рекомбинации в упрощенном виде его можно представить следующим образом:
1. сначала интеграза производит обмен между двумя цепями одинаковой полярности
2. при этом разрыв и воссоединение цепей происходят между строго определенными нуклеотидами в центральной части att-сайтов. В результате возникает структура, физически соответствующая полухиазме Холлидея.
3. затем на расстоянии 7 п.н. происходит вторая пара обменов между двумя другими цепями, не участвовавшими в первом обмене. Вторая пара обменов приводит к интеграции фага; интегрированный профаг фланкирован рекомбинантными сайтами attL и attR, как это уже было показано на рис. 62, в. Предполагается, что для катализа этой реакции необходимы четыре субъединицы интегразы, связанные в att-сайтах.
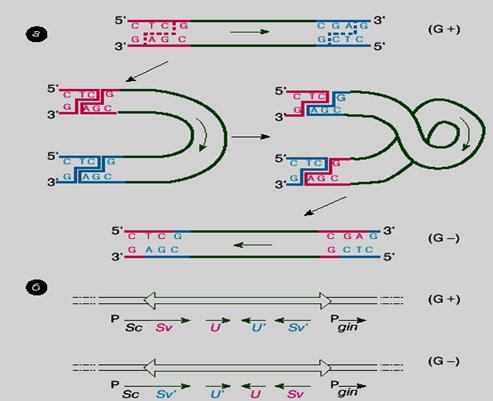
Рисунок. 63. Схема сайт-специфической инверсии сегмента G в ДНК фага Mu (Глазер В.М. 1998).
Сайт-специфическая рекомбинация, катализируемая ферментами второй группы (инвертазы), происходит по более простой схеме (рис. 63), чем в случае с интегразой. Рассмотрим ее на примере инвертазы фага Mu:
1. В центральной части своей хромосомы фаг содержит особый сегмент G размером около 3 т.п.н.
2. Сегмент имеет на концах инвертированные (обращенные) повторы длиной около 30 п.н.
3. В каждом повторе, в свою очередь, имеется специфический рекомбинационный сайт. Инвертаза проводит рекомбинацию между этими сайтами (рис. 63, а).
4. Четыре субъединицы фермента, по одной субъединице на каждую цепь ДНК, делают "временные" разрывы в обеих цепях каждого рекомбинационного сайта, то есть одновременно расщепляют все четыре цепи, образуя 5'-P и 3'-OH-концы. При этом разрывы цепей в каждом дуплексе происходят на расстоянии 2 п.н., так что 3'-конец как бы выступает.
5. Фермент ковалентно связывается с 5'-P-концами.
6. Затем одна часть первого дуплекса меняется местами с такой же частью второго дуплекса, после чего восстанавливаются фосфодиэфирные связи во всех четырех цепях.
| <== предыдущая страница | | | следующая страница ==> |
| МОДЕЛЬ РЕКОМБИНАЦИИ НА ОСНОВЕ РЕПАРАЦИИ ДВУЦЕПОЧЕЧНЫХ РАЗРЫВОВ ДНК | | | Автоматическое повторное включение после АЧР |
Дата добавления: 2014-11-08; просмотров: 717; Нарушение авторских прав

Мы поможем в написании ваших работ!