Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
СТРОЕНИЕ, СВОЙСТВА И ФУНКЦИИ ГЕНОВ
8.1. Ген как дискретная единица наследственности
Одним из фундаментальных понятий генетики на всех этапах ее развития было понятие единицы наследственности. В 1865 году основоположник генетики (науки о наследственности и изменчивости) Г. Мендель на основании результатов своих опытов на горохе пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными единицами наследственности. Единицы наследственности, которые отвечают за развитие отдельных признаков, Г.Мендель назвал «задатками». Мендель утверждал, что в организме по любому признаку имеется пара аллельных задатков (по одному от каждого из родителей), которые между собой не взаимодействуют, не смешиваются и не изменяются. Поэтому, при половом размножении организмов в гаметы попадает лишь один из наследственных задатков в «чистом» неизменном виде.
Позже предположения Г.Менделя о единицах наследственности получили полное цитологическое подтверждение. В 1909 году датский генетик В. Иогансен назвал «наследственные задатки» Менделя генами.
В рамках классической генетики ген рассматривается как функционально неделимая единица наследственного материала, определяющая формирование какого-либо элементарного признака.
Различные варианты состояния определенного гена, возникшие в результате изменений (мутаций), получили название «аллели» (аллельные гены). Количество аллелей гена в популяции может быть значительным, но у конкретного организма число аллелей определенного гена всегда равно двум – по числу гомологичных хромосом. Если в популяции количество аллелей какого-либо гена больше двух, то такое явление получило название «множественного аллелизма».
Гены характеризуются двумя противоположными по биологическому значению свойствами: высокой стабильностью своей структурной организации и способностью к наследственным изменениям (мутациям). Благодаря этим уникальным свойствам обеспечивается: с одной стороны – устойчивость биологических систем (неизменяемость в ряде поколений), а с другой – процесс их исторического развития, формирования адаптаций к условиям окружающей среды, т.е. эволюция.
8.2. Ген как единица генетической информации. Генетический код.
Еще Аристотель более 2500 лет тому высказал предположение о том, что гаметы – это отнюдь не миниатюрные варианты будущего организма, а структуры, содержащие информацию о развитии эмбрионов (хотя он признавал только исключительно важность яйцеклетки в ущерб сперматозоиду). Однако развитие этой идеи в современных исследованиях стало возможным лишь после 1953 года, когда Дж. Уотсон и Ф. Крик разработали трехмерную модель строения ДНК и тем самым создали научные предпосылки для раскрытия молекулярных основ наследственной информации. С этого времени началась эра современной молекулярной генетики.
Развитие молекулярной генетики привело к раскрытию химической природы генетической (наследственной) информации и наполнило конкретным смыслом представление о гене как единици генетической информации.
Генетическая информация – информация о признаках и свойствах живых организмов, заложенная в наследственных структурах ДНК, которая реализуется в онтогенезе через синтез белка. Наследственную информацию, как программу развития организма, каждое новое поколение получает от предков в виде совокупности генов генома. Единицей наследственной информации является ген, который представляет собой функционально неделимый участок ДНК со специфической последовательностью нуклеотидов, определяющей последовательность аминокислот определенного полипептида или нуклеотидов РНК.
Наследственная информация о первичной структуре белка записана в ДНК с помощью генетического кода.
Генетический код – система записи генетической информации в молекуле ДНК (РНК) в виде определенной последовательности нуклеотидов. Этот код служит ключом для перевода последовательности нуклеотидов в и-РНК в последовательность аминокислот полипептидной цепи при ее синтезе.
Свойства генетического кода:
1.Триплетность – каждая аминокислота кодируется последовательностью из трех нуклеотидов (триплетом или кодоном)
2.Вырожденность – большинство аминокислот шифруется более чем одним кодоном (от 2 до 6). В ДНК или РНК имеется 4 различных нуклеотида, которые теоретически могут образовывать 64 разных триплета (4 3 = 64) для кодирования 20 аминокислот, входящих в состав белков. Этим и объясняется вырожденность генетического кода.
3.Неперекрываемость – один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов.
4.Специфичность (однозначность) – каждый триплет кодирует только одну аминокислоту.
5.Код не имеет знаков препинания. Считывание информации с и-РНК при синтезе белка всегда идет в направлении 5, -- 3, в соответствии с последовательностью кодонов иРНК. Если произойдет выпадение одного нуклеотида, то при считывание его место займет ближайший нуклеотид из соседнего кода, из-за чего изменится аминокислотный состав в молекуле белка.
6.Код универсален для всех живых организмов и вирусов: одинаковые триплеты кодируют одинаковые аминокислоты.
Универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов
Однако, универсальность генетического кода не является абсолютной. В митохондриях ряд кодонов имеет другой смысл. Поэтому иногда говорят о квазиуниверсальности генетического кода. Особенности генетического кода митохондрий свидетельствует о возможности его эволюционирования в процессе исторического развития живой природы.
Среди триплетов универсального генетического кода три кодона не кодируют аминокислоты и определяют момент окончания синтеза данной полипептидной молекулы. Это так называемые «nonsens» кодоны (стоп-кодоны или терминаторы). К ним относятся: в ДНК – АТТ, АЦТ, АТЦ; в РНК – УАА, УГА, УАГ.
Соответствие нуклеотидов в молекуле ДНК порядку аминокислот в молекуле полипептида получило название коллинеарности. Экспериментальное подтверждение коллинеарности сыграло решающую роль в расшифровке механизма реализации наследственной информации.
Значение кодонов генетического кода приведены в таблице 8.1.
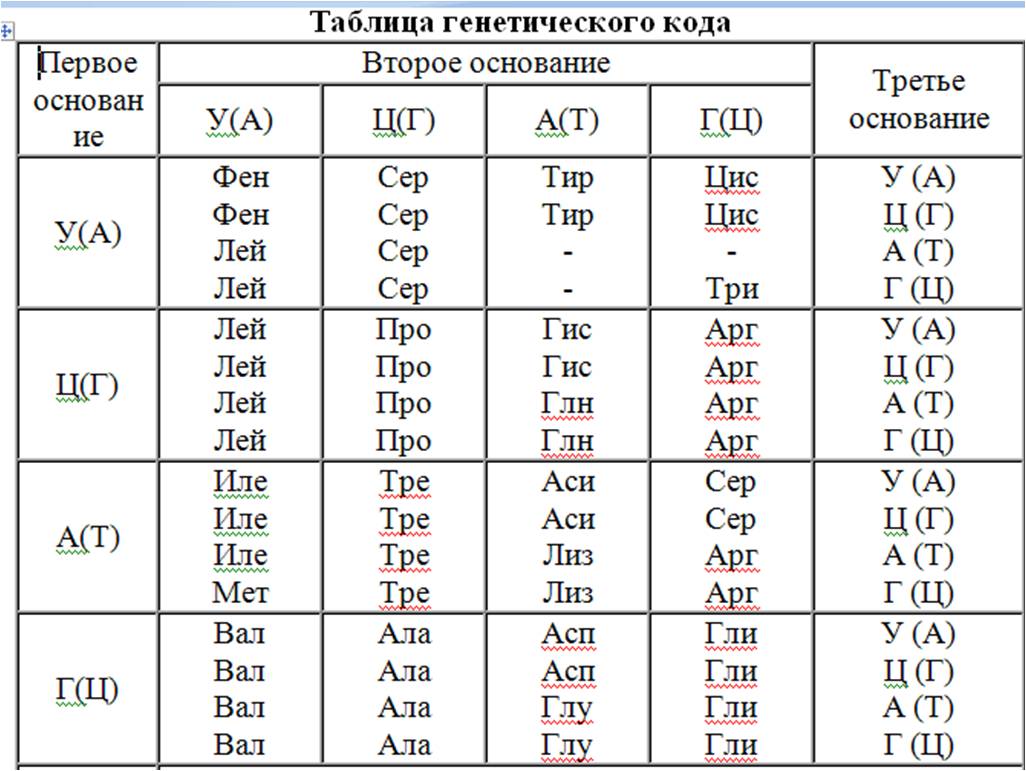
Табл.8.1. Генетический код (кодоны иРНК для аминокислот)
С помощью этой таблицы по кодонам иРНК можно определить аминокислоты. Первый и третий нуклеотиды берут из вертикальных столбиков, расположенных справа и слева, а второй – с горизонтального. В месте перессичения условных линий содержится информация про соответствующую аминокислоту. Отметим, что в таблице приводится триплеты и-РНК, а не ДНК.
Структурно - функциональная организация гена
Молекулярная биология гена
Современное представление о строении и функции гена формировалось в русле нового направления, которое Дж.Уотсон назвал молекулярной биологией гена (1978)
Важным этапом в изучении структурно – функциональной организации гена были работы С. Бензера в конце 1950-хх годов. Они доказали, что ген представляет собой нуклеотидную последовательность, которая может изменятся в результате рекомбинаций и мутаций. Единицу рекомбинации С.Бензер назвал реконом, а еденицу мутации – мутоном. Экспериментально установлено, что мутон и рекон соответствуют одной паре нуклеотидов. Единицу генетической функции С. Бензер назвал цистроном.
В последние годы стало известно, что ген имеет сложное внутренее строение, а отдельные его части обладают разными функциями. В гене можно выделить последовательность нуклеотидов гена, которая определяет строение полипептида. Эта последовательность называется цистроном.
Цистрон – это последовательность нуклеотидов ДНК, которая определяет отдельную генетическую функцию полипептидной цепи. Ген может быть представлен одним или несколькими цистронами. Сложные гены содержащие в себе несколько цистронов называются полицистронными.
Дальнейшее развитие теории гена связано с выявлением различий в организации генетического материала у организмов далеких друг от друга в таксономическом отношении, которыми являются про- и эукариоты.
Структура генов прокариот
У прокариот, типичными представителями которых являются бактерии, большинство генов представлены непрерывными информативными участками ДНК, вся информация которых используется при синтезе полипептида. У бактерий гены занимают 80-90% ДНК. Главная особенность генов прокариот – это их объединение в группы или опероны.
Оперон – это группа следующих подряд структурных генов, находящихся под контролем одного регуляторного участка ДНК. Все сцепленые гены оперона кодируют ферменты одного метаболического пути (например, расщепление лактозы). Такая общая молекула иРНК называется полицистронной. Только некоторые гены прокариот транскрибируются индивидуально. Их РНК называется моноцистронной.
Организация по типу оперона позволяет бактериям быстро переключать метаболизм с одного субстрата на другой. Бактерии не синтезируют ферменты определенного метаболического пути в отсутствии необходимого субстрата, но способны начать их синтезировать при появлении субстрата.
Структура генов эукариот
Большинство генов эукариот (в отличии от генов прокариот) имеют характерную особенность: содержат не только кодирующие структуру полипептида участки – экзоны, но и некодирующие – интроны. Интроны и экзоны чередуются между собой, что придает гену прерывистую (мозаичную) структуру. Количество интронов в генах варьиирует от 2-х до десятков. Роль интронов до конца неясна. Полагают, что они учавствуют в процессах рекомбинации генетического материала, а также в процессах регуляции экспресии (реализации генетической информации) гена.
Благодаря экзонно – интронной организации генов создаются предпосылки для альтернативного сплайсинга. Альтернативний сплайсинг- процесс «вырезания» разных интронов из первичного РНК-транскрипта в результате чего на основе одного гена могут синтезироватся разные белки. Явление альтернативного сплайсинга имеет место у млекопитающих при синтезе различних антител на основе иммуноглобулиновых генов.
Дальнейшие исследование тонкой структуры генетического материала еще больше осложнило четкость определения понятия «ген». В геноме эукариот были обнаружены обширные регуляторные области имеющие различные участки, которые могут располагатся за пределами едениц трансскрипции на расстоянии в десятки тысяч пар нуклеотидов. Структуру эукариотического гена, включающую транскрибируемые и регуляторные области, можно представить следующие образом.
Рис 8.1. Структура эукариотического гена
1 – энхансеры; 2 – сайленсеры; 3 – промотор; 4 – экзоны; 5 – интроны; 6 – участки экзонов, кодирующие нетранслируемые области.
Промотор – участок ДНК для связывания с РНК – полимеразой и образование комплекса ДНК-РНК полимеразы для запуска синтеза РНК.
Энхансеры - усилители транскрипции.
Сайленсеры – ослабители транскрипции.
В настоящее время ген (цистрон) рассматривается как функционально неделимая единица наследственного мастерства, определяющая развитие какого – либо признака или свойства организма. С позиции молекулярной генетики ген представляет собой участок ДНК (у некоторых вирусов РНК), который несет информацию о первичной структуре полипептида, молекулы транспортной и рибосомальной РНК.
В диплоидных клетках человека примерно 32000 пар генов. Большинство генов в каждой клетке «молчит». Набор активных генов зависит от типа ткани, периода развития организма, полученных внешних или внутренних сигналов. Можно сказать, что в каждой клетке «звучит» свой акорд генов, определяя спектр синтезируемых РНК, белков и, соответственно, свойства клетки.
Структура генов вирусов
Вирусы имеют структуру гена, отражающую генетическую структуру клетки - хозяина. Так, гены бактериофагов собраны в опероны и не имеют интронов, а вирусы эукариот имеют интроны.
Характерная особенность вирусных геномов – это явление «перекрывающихся» генов («ген в гене»). В «перекрывающихся» генах каждый нуклеотид принадлежит одному кодону, но имеются разные рамки считывание генетической информации с одной и той же нуклеотидной последовательности. Так, у фага φ Х 174 имеется участок молекулы ДНК, который входит в состав сразу трех генов. Но соответствующие этим генам последовательности нуклеотидов прочитывается каждая в своей системе отсчета. Поэтому нельзя говорить о «перекрывании» кода.
Такая организация генетического материала («ген в гене») расширяет информационные возможности сравнительно небольшого по величине генома вирусов. Функционирование генетического материала вирусов происходит по-разному в зависимости от структуры вируса, но всегда с помощью ферментной системы клетки хозяина. Различные способы организации генов у вирусов, про- и эукариотов представлены на рис 8.2.
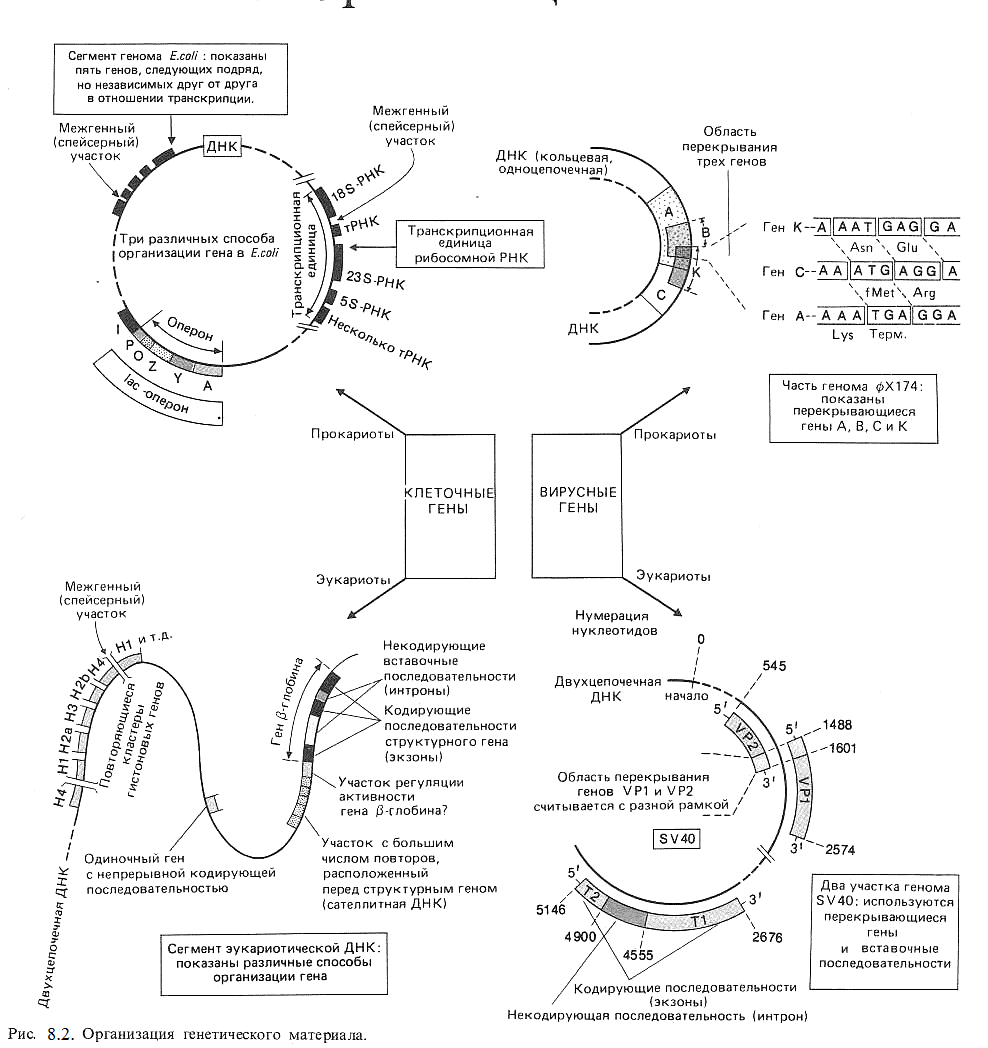
Функционально – генетическая классификация генов
Существует несколько классификаций генов. Так, например, выделяют аллельные и неаллельные гены, летальные и полулетальные, гены «домашнего хозяйства», «гены роскоши» и т.д.
Гены «домашнего хозяйства» - набор активных генов, необходимых для функционирования всех клеток организма независимо от типа ткани, периода развития организма. Эти гены кодируют ферменты транскрипции, синтеза АТФ, репликации, репарации ДНК и др.
Гены «роскоши» имеют избирательную активность. Их функционирование специфично и зависит от типа ткани, периода развития организма, полученных внешних или внутренних сигналов.
Исходя из современных представлений о гене как функционально неделимой единице наследственного материала и системной организации генотипа все гены принципиально можно разделить на две группы: структурные и регуляторные.
Регуляторные гены – кодируют синтез специфических белков, влияющих на функционирование структурных генов таким образом, что в клетках разной тканевой принадлежности синтезируются необходимые белки и в необходимых количествах.
Структурными называются гены, которые несут информацию о первичной структуре белка, рРНК или тРНК. Гены, кодирующие белки, несут информацию о последовательности аминокислот определенных полипептидов. С этих участков ДНК транскребируется иРНК, которая служит матрицей для синтеза первичной структуры белка.
Гены рРНК (выделяют 4 разновидности) содержат информацию о последовательности нуклеотидов рибосомальных РНК и обуславливают их синтез.
Гены тРНК (более 30 разновидностей) несут информацию о строении транспортных РНК.
Структурные гены, функционирование которых тесно связано со специфическими последовательностями в молекуле ДНК, называемыми регуляторными участками, подразделяются на:
· независимые гены;
· повторяющиеся гены;
· кластеры генов.
Независимые гены – это гены, транскрипция которых не связана с транскрипцией других генов в рамках транскрипционной еденицы. Их активность может регулироваться экзогенными веществами, например, гормонами.
Повторяющиеся гены присутствуют в хромосоме в виде повторов одного гена. Ген рибосомной 5-S-РНК повторяется много сотен раз, причем повторы располагаются тандемом, т. е. следуя вплотную друг за другом без промежутков.
Кластеры генов – это локализованные в определенных участках (локусах) хромосомы группы разных структурных генов с родственными функциями. Кластеры тоже часто присутствуют в хромосоме в виде повторов. Например, кластер гистоновых генов повторяется в геноме человека 10-20 раз, обазуя тандемную групу повторов.(рис. 8.3.)
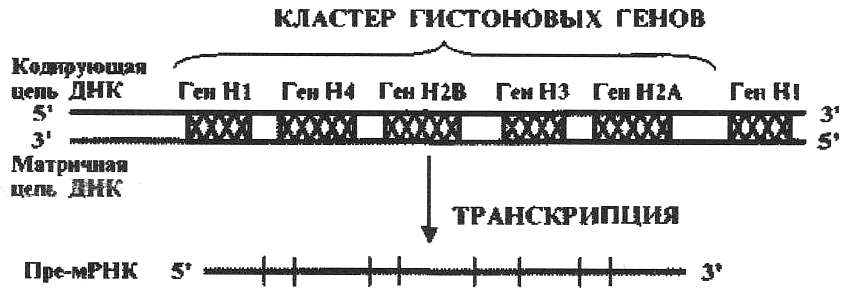
Рис.8.3. Кластер гистоновых генов
За редким исключением кластеры транскрибируются как одно целое – в виде одной длинной пре-мРнк. Так пре-мРНК кластера гистоновых генов содержит информацию про все пять гистоновых белков. Это ускоряет синтез гистоновых белков, которые принимают участие в формировании нуклеосомной структуры хроматина.
Существуют также сложные кластеры генов, которые могут кодировать длинные полипептиды с несколькими ферментативными активностями. Например, один из генов NeuraSpora grassa кодирует полипептид с молекулярной масой 150000 дальтон, который отвечает за 5 последовательных этапов в биосинтезе ароматических аминокислот. Полагают, что полифункциональные белки имеют несколько доменов – конформационно ограниченных полуавтономных образований в полипептидной цепи, выполняющих специфические функции. Открытие полуфункциональных белков дало основание полагать, что они являются одним из механизмов плейотропного действия одного гена на формирование нескольких признаков.
В кодирующей последовательности этих генов могут вклиниваться некодирующие, называемые интронами. Кроме того между генами могут находится участки спейсерной, и сателитной ДНК (рис.8.4).

Рис.8.4. Структурная организация нуклеотидных последовательностей (генов) в ДНК.
Спейсерная ДНК располагается между генами и не всегда транскрибируется. Иногда участок такой ДНК между генами (так называемый спейсер) содержит какую-то информацию, относящуюся к регуляции транскрипции, но он может представлять собой и просто короткие повторяющиеся последовательности избыточной ДНК, роль которой остается неясной.
Сателитная ДНК содержит большое количество групп повторяющихся нуклеотидов, которые не имеют смысла и не транскрибируются. Эта ДНК часто располагается в области гетерохроматина центромер митотических хромосом. Одиночные гены среди сателитной ДНК имеет регулирующие и усиливающие действие на структурные гены.
Большой теоритический и практический интерес для молекулярной биологии и медицинской генетики представляет микро- и минисателитные ДНК.
Микросателитная ДНК – короткие тандемные повторы из 2-6, (чаще из 2-4) нуклеотидов, которые получили название STR. Наиболее распространенными являются нуклеотидные ЦА- повторы. Количество повторов может существенно различатся у разных людей. Микросателиты находятся преимущественно в некоторых участках ДНК и насследуются по законам Менделя. Ребенок получают одну хромосому от матери, с определенным количеством повторов, другую от отца - с другим количеством повторов. Если рядом с геном ответственным за моногенное заболевание, или внутри гена расположен такой кластер микросателитов, то маркером патологического гена может быть определенное количество повторов по длине кластера. Эта особенность используется при непрямой диагностике генных болезней.
Минисателитная ДНК – тандемные повторы из 15-100 нуклеотидов. Они получили название VNTR – вариабельные по количеству тандемные повторы. Длина этих локусов также существено вариабельна у разных людей и может быть маркером (меткой) патологического гена.
Микро- и макросателитные ДНК используют:
1. Для диагностики генных болезней;
2. В судебно-медицинской экспертизе для идентификации личностей;
3. Для установления отцовства и в других ситуациях.
Наряду со структурными и регуляторными повторяющимися последовательностями, функции которых неизвестны, обнаружены мигрирующие нуклеотидные последовательности (транспозоны, мобильные гены), а также так называемые псевдогены у эукариот.
Псевдогены – нефункционирующие последовательности ДНК, которые сходные с функционирующими генами.
Вероятно, они произошли путем дупликации, а неактиаными копии стали в результате мутаций, нарушивших какие-либо стадии экспрессии.
По одной из версий псевдогены являются «эволюционным резервом»; по-другой – представляют собой «тупики эволюции», побочный эффект перестроек некогда функционирующих генов.
Транспозоны — структурно и генетически дискретные фрагменты ДНК, способные перемещаться от одной молекулы ДНК к другой. Впервые предсказаны Б.Мак-Клинток (рис. 8) в конце 40-х годов XX века на основе генетических экспериментов на кукурузе. Изучая природу окраски зерен кукурузы она сделала предположение, что существуют так званые мобильные („прыгающее”) гены, которые могут перемещаться по геному клетки. Пребывая по соседству с геном ответственным за пигментацию зерен кукурузы мобильные гены блокируют его работу. В дальнейшем транспозоны были выявлены у бактерий и было установлено, что они ответственны за устойчивость бактерий к различным токсическим соединениям.

Рис. 8.5. Барбара Мак- Клинток Впервые предсказала о существовании мобильных («прыгающих») генов, способных перемещаться по геному клеток.
Мобильные генетические элементы выполняют такие функции:
1. кодируют белки, ответственные за их перемещение и репликацию.
2. вызывают многие наследственные изменения в клетках, вследствии чего образуется новый генетический материал.
3. приводит к образованию раковых клеток.
4. встраиваясь в различные участки хромосом, они инактивируют или усиливают экспрессию клеточных генов,
5. является важным фактором биологической эволюции.
Современное состояние теории гена
Современные теории гена сформирована благодаря переходу генетики на молекулярный уровень анализа и отражает тонкую структурно-функциональную организацию единиц насследственности. Основные положения этой теории следующие:
1) ген(цистрон) – функциональная неделимая еденица наследственного материала (ДНК у организмов и РНК у некоторых вирусов), определяющая проявление наследственного признака или свойства организма.
2) Большинство генов существует в виде двух или большего числа альтернативных (взаимоисключающих) вариантов аллелей. Все аллели данного гена локализуются в одной и той же хромосоме в определенном ее участке, которую назвали локусом.
3) Внутри гена могут происходить изменения в виде мутаций и рекомбинаций; минимальные размеры мутона и рекона равны одной паре нуклеотидов.
4) Существуют структурные и регуляторные гены.
5) Структурные гены несут информацию о последовательности аминокислот в определенном полипептиде и нуклеотидов в рРНК, тРНК
6) Регуляторные гены контролируют и направляют роботу структурных генов.
7) Ген не принимает непосредственного участия в синтезе белка, он является матрицей для синтеза различных видов РНК, которые непосредственно принимают участие в синтезе белка.
8) Существует соответствие (колиннеарность) между расположением триплетов из нуклеотидов в структурных генах и порядком аминокислот в молекуле полипептида.
9) Большинство мутаций гена не проявляются в фенотипе, так как молекулы ДНК способны к репарации (востановлению своей нативной структуры)
10) Генотип являет собой систему, которая состоит из дискретных едениц – геннов.
11) Фенотическое проявление гена зависит от генотипической среды, в которой находится ген, влияние факторов внешней и внутренней среды.
| <== предыдущая страница | | | следующая страница ==> |
| ОБМЕН ВЕЩЕСТВ (МЕТАБОЛИЗМ) И ЭНЕРГИИ В КЛЕТКЕ КЛЕТОЧНЫЕ МЕМБРАНЫ, ИХ СТРОЕНИЕ И ФУНКЦИИ | | |
Дата добавления: 2014-10-02; просмотров: 1450; Нарушение авторских прав

Мы поможем в написании ваших работ!