Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Зависимость скорости энергетического обмена от массы тела особей
Поскольку энергетические траты организма можно определить по количеству потребленного им кислорода, такие термины, как «скорость дыхания», «скорость энергетического обмена» и «скорость метаболизма» часто используются как синонимы.
Скорость дыхания особи, или количество кислорода, потребленное за единицу времени (Q), возрастает с увеличением массы ее тела (W). При этом Q возрастает в значительно меньшей степени, чем W. Это положение хорошо иллюстрируют данные по скорости дыхания у млекопитающих разных размеров (табл. 3.1). Из них следует, что при возрастании массы тела особей в 80 тыс. раз, скорость их дыхания увеличивается только в 7,55 тыс. раз.
Таблица 3.1. Скорость и интенсивность потребления кислорода у млекопитающих с разной массой тела (по Шмидт-Ниельсену, 1982).
| № | Вид животных | Масса, кг | Q, лО2 ·особь-1·час-1 | Q/W, лО2 ·кг-1·час-1 |
| Землеройка | 0,0048 | 0,0355 | 7,40 | |
| Мышь-малютка | 0,0090 | 0,0225 | 2,5 | |
| Малый прыгун | 0,0152 | 0,0273 | 1,80 | |
| Мышь | 0,025 | 0,041 | 1,65 | |
| Суслик | 0,096 | 0,09 | 1,02 | |
| Крыса | 0,290 | 0,25 | 0,87 | |
| Кошка | 2,5 | 1,70 | 0,68 | |
| Собака | 11,7 | 3,87 | 0,33 | |
| Овца | 42,7 | 9,59 | 0,22 | |
| Человек | 14,76 | 0,21 | ||
| Лошадь | 71,1 | 0,11 | ||
| Слон | 268,0 | 0,07 |
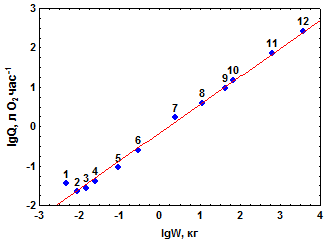
Эти же данные, отложенные в двойных логарифмических координатах (lgQ – lgW), удовлетворительно располагаются вдоль прямой линии (рис. 3.1а). Отсюда следует, что зависимость Qот Wу исследованной группы млекопитающих в общем виде следует степенному уравнению:
Q = aWb, (3.1)
где а – эмпирический коэффициент («уровень метаболизма»), численно равный Qпри W= 1; b– степенной коэффициент, который может изменяться от 0,55 до 1, однако обычно находится в пределах 0,65–0,90.
В численной форме уравнение (3.1) для исследованной группы млекопитающих, рассчитанное по данным табл. 3.1, имеет вид
Q = 0,70W0,711, (3.2)
где Q– скорость дыхания, лО2·особь-1·час-1; W– масса тела, кг.
| 
|
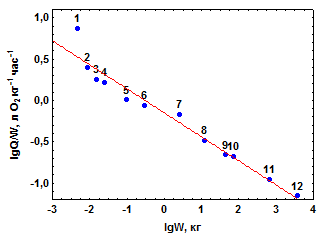
| Рис. 3.1.Зависимость скорости (а) и интенсивности (б) дыхания у млекопитающих от массы их тела. Прямая на рис 3.1а – линия регрессии уравнения (3.2), на рис 3.1б – уравнения (3.5). Номера точек соответствуют порядковым номерам видов в таблице 3.1 |
Степенной характер зависимости Qот Wнаблюдается не только при межвидовых, но и при внутривидовых сопоставлениях, если диапазон изменения значений Wу исследованных особей достаточно широк – не менее 1–1,5 порядков. При логарифмировании уравнение (3.1) трансформируется в уравнение прямолинейной регрессии:
lgQ = lga + b lgW. (3.3)
На графике lgQ – lgW (рис. 3.1б) lga характеризует смещение по вертикали положения линии регрессии lgQ по lgW относительно пересечения осей координат, тогда как коэффициент bравен тангенсу угла наклона линии регрессии к оси абсцисс.
Часто считают, что чем больше коэффициент а, тем выше уровень метаболизма того или иного вида. Однако значения коэффициентов а и b в уравнении (3.1) зависят друг от друга и от размерности величин W (кг, г, мг и т.д.) и Q(млО2·особь-1час-1, гО2·особь-1сутки-1 и т.д.). Поэтому сопоставление уровней метаболизма у разных видов или групп животных по коэффициентам aправомерно лишь в случаях, когда значения Q и Wу них выражены в одних единицах, а значения коэффициентов b очень близки.
Хотя скорость дыхания млекопитающих возрастает с увеличением массы их тела, интенсивность их дыхания (Q/W),иликоличество кислорода, потребленного в расчете на единицу их массы за единицу времени, напротив, снижается (табл. 3.1). Эта связь также может быть описана степенным уравнением:
Q/W =(aWb)/W= aW(b-1). (3.4)
Поскольку значения степенного коэффициента b в (3.1) изменяются в пределах от 0,55 до 1, то b–1варьируют от -0,45 до 0, но обычно в пределах от -0,35 до -0,10. Например, в уравнении (3.4) для упомянутой уже группы млекопитающих (табл. 3.1) значение b = -0,729, поскольку, исходя из (3.1)
Q/W = 70W(1–0,711) = 70W(– 0,289).(3.5)
Так как b–1 < 0, уравнение (3.4) в двойных логарифмических координатах трансформируется в уравнение нисходящей прямолинейной регрессии (рис. 3.1б).
По уравнениям типа (3.1) возможно рассчитать метаболические траты особи, если известна масса ее тела. Например, данное уравнение для большой группы хрящевых и костных рыб при 20 оС имеет вид
Q = 0,223W0,84, (3.6)
где Q –млО2·особь-1·час-1, W– сырая масса особей (г) (Зотин, Зотина, 1999).
Отсюда суточные траты энергии на дыхание (Т) у рыбы массой 1 кг составляют: Т = 0,223(1000)0,84 –24 часа– 4,86 кал·млО2-1 = 8,613 ккал·сутки-1, или 36,037 кДж·сутки-1.
В последнее время скорость метаболизма организмов часто выражают не в единицах потребленного ими кислорода или соответствующей этому энергии, а в единицах мощности, т.е. в ваттах (напр., Зотин, Зотина, 1999; Дольник, 2002).Поскольку 1 ватт (Вт) = 1 Дж·сек-1, коэффициенты пересчета значений скоростей энергетического метаболизма в единицы мощности составляют 1 ккал·особь-1·час-1 = 1,1622 Вт, или 1 млО2·особь-1час-1 = 0,00558 Вт.
Отсюда рассчитанная по (3.6) скорость метаболизма рыбы массой 1 кг, равная 73,8 млО2·особь-1·час-1, соответствует мощности 73,8 млО2 особь-1час-1 • 4,86 кал млО2-1· 4,184 Дж·кал-1 /3600 секунд = 0,417 Вт·особь-1.
Из уравнения (3.4) следует, что интенсивность метаболизма у крупных организмов ниже, чем у мелких. Поэтому последние способны развивать более высокую мощность в расчете на единицу массы своего тела. Например, муравей может передвигать предметы весом в несколько раз больше его собственного веса, тогда как слон способен переносить груз не более половины собственного веса. Аналогичная зависимость имеет место и у человека; ее наиболее наглядным примером являются рекорды у тяжелоатлетов. Например, на 1 мая 2005 г. мировой рекорд в толчке у мужчин в весовой категории до 56 кг составлял 168 кг, что в три раза больше массы атлета. В то же время, у супертяжеловесов, масса которых в среднем достигает 130 кг, мировой рекорд равен 263,5 кг, т. е. только в два раза выше их массы.
3.3. Зависимость скорости метаболизма у животных от температуры.На скорость дыхания организмов, как и на другие их жизненные процессы, значительное воздействие оказывает температура среды. Поскольку масса особей также влияет на скорость дыхания, воздействие температуры на этот показатель следует изучать на одноразмерных особях.
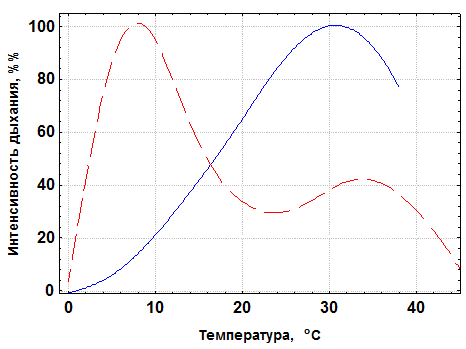
Характер температурной зависимости скорости (Q) или интенсивности дыхания (Q/W) у пойкилотермных и гомойотермных животных резко различается (рис. 3.2).
|
| Рис. 3.2.Зависимость интенсивности дыхания от температуры у пойкилотермных (сплошная линия) и гомойтермных (пунктир) животных (схема). За 100 % у обеих групп принята максимальная интенсивность дыхания |
У пойкилотермных животных,температура тела которых равна или очень мало отличается от температуры среды, значения Q или Q/W с температурой возрастают, а Q10 для них снижаются. Примером этого являются приводимые уже данные для черноморского моллюска Modiola adriatica (рис. 1.2). При изучении изменений Q/Wу колорадского жука в широком температурном диапазоне выявлена аналогичная закономерность (табл. 3.2).
Заметим, что значения ΔFдля Q/Wуэтого вида (см. гл. 1) остаются практически постоянными в интервале 7–20 оС, но быстро снижаются при дальнейшем повышении температуры. Следовательно, температурная зависимость Q/W у колорадского жука может быть описана уравнением Вант-Гоффа – Аррениуса только в температурном диапазоне 7–20 оС.
Однако возрастание Q (или Q/W) с температурой у пойкилотермных животных имеет место лишь в пределах оптимальных температур, понимаемых здесь в широком смысле, включая зоны оптимума и нормы на кривой толерантности Шелфорда. При дальнейшем повышении температуры за пределы этой зоны скорость метаболизма снижается. Проиллюстрируем это на примере эвритермного легочного моллюска Lymnaea hoduthae (Хмелева, Голубев, Лаенко, 1985). Данный вид обитает в геотермальном источнике Ходутка (Камчатка) в интервале температур от 5–7 до 35 оС, но не теряет жизнеспособности в течение нескольких часов при ее повышении до 40 оС, однако размножается в более узком интервале – от 12 до 33 оС
Таблица 3.2. Интенсивность дыхания у колорадского жука Leptonotarsa decemlineata при разных температурах (по Marzusch, 1952)
| Температура, оС | Интенсивность дыхания, мклО2 г-1·час-1 | Температурный интервал, оС | Q10 | ∆F |
| 7 – 10 | 2,57 | 14 855 | ||
| 10 – 15 | 2,41 | 14 238 | ||
| 15 – 20 | 2,52 | 15 490 | ||
| 20 – 25 | 2,10 | 12 866 | ||
| 25 – 30 | 1,56 | 7 974 |
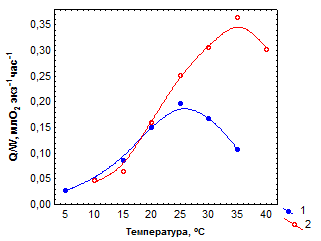
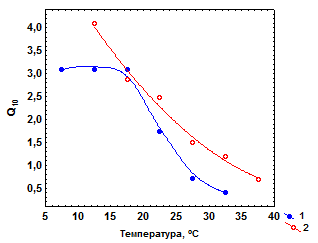
Исследована температурная зависимость Q/Wу особей из двух субпопуляций L. hoduthae, обитающих в зонах источника с градиентами температур 8–40 оС и 25–33 оС. Значения Q/W при равных температурах в верхней части температурной шкалы зависят от температурных условий обитания особей. Так, их максимальные показатели у моллюсков из первой субпопуляции отмечены при 25о С, а из второй – при 35 оС (рис. 3.3а).
| 
|
| Рис. 3.3.Зависимость интенсивности дыхания (Q/W) и Q10 для этого показателя от температуры у одноразмерных особей из двух субпопуляций Lymnaea hodutkae, обитающих в зонах геотермального источника с градиентами температур 8–40 оС (1) и 25–33 оС (2) |
Тем не менее, характер изменений Q/W с температурой в обоих случаях одинаков. Этот показатель вначале возрастает с температурой, достигает максимума при определенных ее значениях вблизи верхнего предела зоны температурного оптимума, а затем резко снижается. Значения Q10 для Q/Wв целом снижаются с температурой, становясь в сублетальной зоне меньше единицы (рис. 3.3б).
Снижение Q/Wпри повышенных температурах часто сопровождается ростом смертности особей. Поэтому характер температурной зависимости Q/Wможет являться одним из критериев нахождения верхней температурной границы зоны нормы.
У отдельных видов пойкилотермных животных значения коэффициента а в уравнении (3.1) возрастают с температурой в пределах толерантных ее значений, при этом величины Q10 составляют 1,5–2,5, Напротив, степенной коэффициент b с ростом температуры обычно снижается (табл. 3.3).
Таблица 3.3. Значения коэффициентов а и b уравнения (3.1) зависимости скорости дыхания (Q, млО2·экз-1·час-1) от сырой массы тела (W, г) у дальневосточного трепанга Stichopus japonicus (по Рябушко, 2000)
| Температура, оС | a | b |
| 2,5 | 0,0029 | 0,745 |
| 8,5 | 0,0070 | 0,85 |
| 13,5 | 0,025 | 0,65 |
| 18,5 | 0,11 | 0,38 |
| 23,5 | 0,12 | 0,41 |
| 28,5 | 0,32 | 0,29 |
У гомойотермных животных (рис. 3.2) на характер температурной зависимости скорости метаболизма значительное воздействие оказывает необходимость поддержания постоянной температуры тела при изменениях температуры среды. Если температура их тела выше температуры среды (а чаще всего так и бывает), часть образующегося в процессе метаболизма тепла рассеивается в среде. Согласно закону остывания Ньютона, величина теплоотдачи (Н) сплошного нагретого тела (шара) равна произведению удельной теплопроводности тела (C), площади его поверхности (S) и разности температуры поверхности тела (Тт) и среды (Тс), т. е.
Н = CS(Tт –Тс).(3.7)
Ряд ученых, начиная с М. Рубнера (1882), считает возможным использовать уравнение (3.7) для характеристики величин потерь метаболического тепла у гомойотермных животных. В частности, оно вполне подходит для человека, у которого очень слабо развиты волосяной покров и подкожная жировая прослойка, выполняющие теплоизолирующую функцию.
Однако для большинства птиц и млекопитающих, имеющих хорошо развитые внешние покровы (мех, перья и т. п.) или толстый слой подкожного жира, более подходит закон Фурье, описывающий теплоотдачу тел, покрытых теплоизолирующей оболочкой:
Н = hS(Tт–Тс)/L,(3.8)
где h – удельная теплопроводность внешних покровов, L– их толщина, S – площадь поверхности тела (Дольник, 2002).
Из (3.7) и (3.8) следует, что чем выше разность Tт–Тс, тем большее количество тепла (при прочих равных условиях) передается от нагретого тела во внешнюю среду. Величина Тс у отдельных видов достаточно постоянна; для всей поверхности тела человека, например, она составляет в среднем около 25-27оС. Поэтому чем ниже температура среды, чем больше тепла теряет организм, тем выше должен быть уровень его метаболизма для компенсации потерь тепла.
Поэтому максимальный уровень метаболизма у гомойотермных животных («вершинный обмен») отмечается при некоторой температуре, близкой к нижнему пределу зоны ее толерантных значений («температура холодового оцепенения»). При дальнейшем повышении температуры скорость метаболизма снижается, достигая некоторого минимального значения («основной обмен») при определенной температуре среды («критическая точка») или в узком температурном интервале («термонейтральная зона»). «Критическая точка» равна или очень близка к температуре внешних покровов животного, поэтому при ней между организмом и средой устанавливается тепловое равновесие и теплоотдача организма во внешнюю среду становится равной нулю. Например, у человека минимальный уровень обмена наблюдается в узком температурном интервале 25–27 оС, практически совпадающим со средней температурой поверхности его тела.
При температуре среды выше критической точки тепло из внешней среды поступает в организм, что ведет к его перегреву. Это делает необходимым отведение излишнего тепла из организма, что достигается посредством усиленного потоотделения, учащения дыхания, увеличения поверхностного кровотока. На эти процессы расходуется дополнительная энергия, что приводит к некоторому повышению уровню метаболизма. При дальнейшем повышении температуры уровень метаболизма быстро снижается, становясь равным нулю при температуре смерти от перегрева (Слоним, 1971).
3.4. Уровни метаболизма у важнейших таксономических групп животных.Сравнительная оценка уровней метаболизма у видов из разных таксонов имеет большое теоретическое значение для оценки сдвигов уровня метаболизма в процессе прогрессивной эволюции. А. Хеммингсен (1960) впервые обобщил данные по скорости дыхания у разных групп гетеротрофных пойкилотермных и гомойотермных организмов. Он установил, что зависимость скорости дыхания (Q) от массы тела у гетеротрофных протистов (в основном саркодовых), пойкилотермных (беспозвоночных и позвоночных) и гомойотермных (млекопитающих и птиц) животных соответствует уравнениям типа (3.2) со степенными коэффициентами b, очень близкими к 0,75. Это дало возможность сопоставить уровни метаболизма у этих групп по величинам коэффициентов а.
На основании этого А. Хеммингсен пришел к выводу, что средний уровень обмена у исследованных пойкилотермных животных приблизительно на порядок превосходил уровень обмена у протистов и на столько же уступал таковому для гомойотермных животных. Отсюда был сделан вывод о скачкообразном повышении уровней метаболизма при ароморфозах, приведших в процессе прогрессивной эволюции к появлению вначале многоклеточных, а затем и гомойотермных животных. Эти взгляды получили широкое распространение. Уравнения, рассчитанные А. Хеммингсеном и построенный им график зависимости lgQ – lgWдля указанных групп организмов, получили широкую известность и на протяжении нескольких десятков лет включались в учебники экологии.
Однако обобщения последних лет, основанные на анализе значительно большего фактического материала (табл. 3.4), внесли значительные коррективы в эти представления. Оказалось, что уровень метаболизма у низкоорганизованных протистов из типа саркодовых лишь незначительно уступает таковому у большинства групп беспозвоночных, тогда как уровень метаболизма у инфузорий (самых высокоорганизованных протистов) даже более высокий, чем у позвоночных пойкилотермных животных. В то же время уровень обмена у самых мелких многоклеточных животных – коловраток ниже, чем у саркодовых.
Наивысший уровень метаболизма среди пойкилотермных животных имеют активно летающие насекомые из отрядов стрекоз, чешуекрылых, перепончатокрылых и двукрылых. По этому показателю они в 1,5–4 раза превосходят нелетающих или плохо летающих насекомых из отрядов коллембол, прямокрылых и жесткокрылых.
Даже выход предков амфибий на сушу, т. е. попадание их под воздействие земного тяготения, а затем появление настоящих наземных позвоночных – рептилий, не привело к существенному повышению уровня обмена этих групп по сравнению с их водными предковыми формами – рыбами.
В целом скорость дыхания у пойкилотермных позвоночных значительно ниже, чем у равных им по массе гомойотермных животных даже с учетом существенной разницы в температуре тела – соответственно около 20 и 40 оС. Пересчитанные для 40 оС значения aв уравнениях (3.1), представленных в табл. 3.3, для рыб, амфибий и рептилий, рассчитанные для температур около 20 оС исходя из Q10 = 2,3, не превышают 10–12, что гораздо ниже аналогичных показателей в соответствующих уравнениях для птиц и млекопитающих.
Очевидно, скачкообразные повышения (в десятки раз) уровня метаболизма в эволюции животных имели место лишь при возникновении млекопитающих, а затем и птиц от их предков-рептилий. При этом уровень обмена у сумчатых млекопитающих несколько уступает таковому у плацентарных, что вполне объясняется более высокой температурой тела у последних.
Таблица 3.4.Параметры обобщенных уравнений типа Т = aWk связи скорости метаболизма (Т, ккал·особь-1сутки-1) от массы тела особей (W, кг) для важнейших таксонов протистов, беспозвоночных и позвоночных животных
| Таксон* | Температура тела, оС | а | k |
| Саркодовые | Около 20 оС | 1.06 | 0.75 |
| Свободноживущие инфузории | -"- | 9,26 | 0.75 |
| Коловратки | -"- | 0,40 | 0,72 |
| Нелетающие насекомые | -"- | 3,20 – 7,46 | 0,76 – 0,85 |
| Летающие насекомые | -"- | 8,00 – 17,30 | 0,82 – 0,91 |
| Ракообразные | -"- | 2,80 | 0,76 |
| Хрящевые и костные рыбы | -"- | 4,63 | 0,84 |
| Амфибии | -"- | 2,02 | 0,81 |
| Рептилии | -"- | 2.20 | 0,79 |
| Птицы из отряда воробьинообразных | 41–42 оС | 114,8 | 0,73 |
| Птицы из остальных отрядов | 41–42 оС | 73,5 | 0,73 |
| Сумчатые млекопитающие | 33–34 оС | 46,6 | 0,74 |
| Плацентарныемлекопитающие | 37–39 оС | 70,0 | 0,75 |
| * уравнения для саркодовых и инфузорий рассчитаны по данным В.В.Хлебовича (1987), коловраток – по Г.А.Галковской и Г.Г.Винбергу (1979), ракообразных – по Л.М.Сущене (1972), для остальных таксонов – по В.Р.Дольнику (2002). |
В то же время уровень метаболизма у птиц из отряда воробьинообразных в 1,5 раза выше, чем плацентарных млекопитающих, а также у птиц из остальных отрядов, хотя температура тела у видов из этих групп очень близка. Причины повышенного уровня обмена у воробьинообразных по сравнению с другими гомойотермными животными остаются неизвестными.
| <== предыдущая страница | | | следующая страница ==> |
| Энергетический обмен организмов | | | Активный обмен у животных |
Дата добавления: 2014-11-15; просмотров: 744; Нарушение авторских прав

Мы поможем в написании ваших работ!