Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
ПРЕДЕЛ ЧИСЛЕННОСТИ ВИДА
B d
Рис. 3.6. Возрастная пирамида популяции полевки
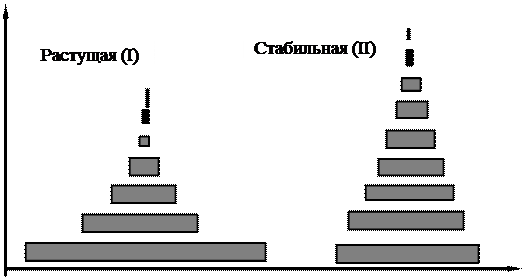
Рис. 3.5. Типы возрастных пирамид
Рис. 3.4. Разные типы кривых выживания
Рождаемость (плодовитость) - характеризует процесс появления на свет новых особей любого организма независимо от того, рождаются ли они, вылупляются из яиц, прорастают из семян или появляются в результате деления, что приводит к увеличению численности популяции. Этот термин используется также и при анализе популяций человека (в демографии). Обычно рождаемость выражают либо как скорость, определяемую путем деления общего числа вновь появившихся особей на время (общая рождаемость), либо как число вновь появившихся особей в единицу времени на 1 особь в популяции (удельная рождаемость).
В живых организмах заложена огромная потенциальная возможность к размножению. Подсчитано, что бактерии делятся каждые 20 минут. При таком темпе одна клетка за 36 часов дает потомство, которое может покрыть сплошным слоем всю планету. Один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут. В действительности же такая громадная плодовитость организмов никогда не реализуется. В связи с этим различают максимальную или потенциальную рождаемость и экологическую или реализованную рождаемость.
Потенциальная рождаемость - это теоретически возможная максимальная скорость образования новых особей в идеальных условиях, когда отсутствует действие лимитирующих экологических факторов, а размножение сдерживается только физиологическими возможностями организма.
Экологическая или реализованная рождаемость означает увеличение численности популяции при фактических условиях среды. Эта величина не постоянна и изменяется в зависимости от размерного и возрастного состава популяции и физических условий среды.
Например, в колонии чайки отложено 510 яиц (потенциальная рождаемость) из них вылупилось 265 птенцов, реализованная плодовитость составила 52% от потенциальной. Мучные хрущики отложили 1200 яиц – вылупилось 713 особей, что составило всего 60%. У капустной белянки в результате гибели яиц, гусениц и куколок по разным причинам (болезни, паразиты, хищники и др.) на свет появляется лишь 0,32 % взрослых бабочек от числа отложенных яиц.
Рождаемость каждого вида определяется как приспособление, обеспечивающее пополнение популяций, а значит, ее жизнестойкость. Поэтому естественно, что у менее приспособленных к неблагоприятным условиям видов высокая смертность в молодом возрасте компенсируется значительной плодовитостью. В зависимости от взаимоотношений между родителями и потомками существует две противоположные стратегии размножения. Виды животных и растений, у которых забота о потомстве выражена явно - смертность их потомства на ранних стадиях развития низка, а сами они имеют низкую плодовитость. Потомство видов, у которых отсутствует забота о детенышах, имеют высокую смертность, а их родители - высокий репродуктивный потенциал (как компенсацию за высокую смертность потомков). Для видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая потенциальная и низкая экологическая рождаемость. В благоприятных условиях рождаемость обычно низкая а в неблагоприятных – высокая. Этим объясняется более высокая рождаемость животных в зонах умеренного климата по сравнению с тропическими видами.
Смертность. Численность и плотность популяции зависят также от ее смертности. Смертность популяции - это количество особей, погибших за определенный период. Она, как и рождаемость, изменяется в зависимости от условий среды, возраста и состояния популяции и выражается в процентах к начальной или чаще к средней ее величине. Выделяют: экологическую смертность - гибель особей, наблюдаемую в конкретных условиях среды (эта величина непостоянна и изменяется в зависимости от условий среды и состояния самой популяции); теоретическую минимальную смертность - величина, постоянная для популяции; она представляет собой гибель особей в идеальных условиях, при которых популяция не подвергается действию лимитирующих факторов. Даже в самых лучших условиях особи будут умирать от старости. Этот возраст определяется физиологической продолжительностью жизни, которая часто превышает среднюю экологическую продолжительность жизни.
Различают четыре основных типа возрастных изменений смертности (рис. 3.4).
I. Первый тип смертности характеризуется повышенной гибелью взрослых, главным образом старых особей. Наблюдается он у многих видов насекомых, личинки которых обитают в почве, воде, древесине или других местах с благоприятными условиями.
II. Ступенчатая кривая (выживание резко меняется при переходе от одной стадии жизненного цикла к другой).
 |
III. К третьему относится животные и растения, смертность которых одинакова во всех возрастах. Она описывается уравнением прямой. Такая смертность встречается очень редко и только у популяций, постоянно находящихся в оптимальных условиях.
IV. Четвертый тип смертности отличается повышенной гибелью особей на ранних стадиях развития и свойствен большинству растений и животных. Максимальная гибель многих растений происходит в стадии прорастания семян и всходов, а животных - в личиночной фазе или в молодом возрасте (у многих рыб до взрослой фазы доживает 1- 2 % от числа выметанной икры, у насекомых - 0,3-0,5 % от отложенных яиц).
Чаще всего процессы смертности оцениваются по кривым выживания. Они выражают зависимость числа выживших из 100 или 1000 особей от их возраста.
Форма кривой выживания связана со степенью заботы о потомстве и другими способами защиты молоди. Так, кривые выживания пчел и дроздов (которые заботятся о потомстве) значительно менее вогнуты, чем у кузнечиков и сардин (которые не заботятся о потомстве). У этих видов это обстоятельство компенсируется значительно большим числом откладываемых яиц.
Форма кривой выживания очень часто изменяется так же в зависимости от плотности популяции. На мало заселенной территории больше шансов выжить. То же самое можно сказать и относительно человеческих популяций, где высокая плотность не всегда бывает благоприятна для отдельного индивидуума. Многие экологи считают, что быстрый рост и высокая плотность в популяциях человека опасны не столько для выживания, сколько для качества жизни индивидуума. И это несмотря на то, что экологическая продолжительность жизни человека значительно увеличилась благодаря достижению современной медицины, улучшению питания и т. д. Однако физиологическая продолжительность жизни человека, по-видимому, не увеличилась, поскольку в наше время до 100 лет доживает не больше людей, чем в минувшие столетия.
На динамику численности популяции большое влияние оказывают миграции отдельных особей, т. е. обмен индивидуумов популяций. Это происходит при переселении их из одного местообитания в другое. В основном миграции совершаются при подрастании молодняка и его расселении. В результате миграций устраняется избыток особей в одной популяции и компенсируется недостаток в другой.
Возрастная структура популяции. Возрастная структура популяции является ее важной характеристикой, которая влияет как на рождаемость, так и на смертность. Соотношение разных возрастных групп в популяции определяет ее способность к размножению в данный момент и показывает, что можно ожидать в будущем. У большинства видов репродуктивная способность их членов изменяется с возрастом. В начальный период (который может продолжаться всего несколько минут, как у микроорганизмов, или несколько лет, как у человека и многих видов деревьев) организмы размножаться не способны. За этим периодом следует период постепенного повышения способности к размножению, достигающей максимума, а затем начинается спад, доходящий, в конечном счете, до полного прекращения. Соответственно в популяции обычно выделяют три экологические возрастные группы (фазы): 1). предрепродуктивную; 2). репродуктивную; 3). пострепродуктивную (рис. 3.5). Длительность этих возрастных фаз по отношению к общей продолжительности жизни варьирует у разных организмов. У современного человека три этих возраста приблизительно одинаковы, на каждый из них приходится около трети жизни (рис.3.7). У первобытных людей пострепродуктивный период был намного короче. Для многих животных и растений характерен очень длительный предрепродуктивный период. В большинстве природных популяций особи гибнут, не достигнув пострепродуктивного периода. Однако люди, домашние животные, а также животные, которых содержат в зоопарках, живут в более благоприятных условиях, а поэтому длительность пострепродуктивного периода может быть значительной. Обычно в быстро растущих популяциях значительную долю составляют молодые особи (рис. 3.5 (I), рис. 3.6 (I)) в популяции, находящейся в стационарном состоянии (в стабильных популяциях), возрастное распределение более равномерно (рис. 3.5 (II), рис. 3.6 (II)), а в популяции, численность которой снижается, будет содержаться большая доля старых особей (рис. 3.5, (III)). Возрастная структура популяции может изменяться и без изменения ее численности. Любая популяция, находящаяся в стабильной среде при благоприятных условиях, в конце концов, приближается к некоторому стабильному возрастному распределению.
 |  | | |||






 I II III
I II III
 | |||||||
 | |||||||
 | |||||||
 | |||||||


|
Временные отклонения от него могут быть вызваны лишь необычными увеличениями рождаемости или смертности, после чего происходит возврат к стабильной ситуации.
 | |||||
 | |||||
| |||||
 |
Рис. 3.7. Возрастная структура популяции человека, рассчитанная отдельно для мужчин и женщин в Швеции и Коста – Рике. Население Швеции увеличивается медленно, поэтому основания пирамиды узкое. Процент старших возрастных классов (40 - 60 лет) велик. Быстрый рост населения Коста – Рики сформировал пирамиду с широким основанием и высоким процентом населения моложе 25 лет (Ricklefs, 1979).
Изучение половозрастного состава популяции имеет большое значение. Один из основных факторов, влияющих на размеры популяции, - это процент особей, погибающих до достижения половой зрелости (смертность в дорепродуктивном периоде). В пределах вида эта величина гораздо более изменчива, чем рождаемость. У промысловых птиц, и пушных зверей отношение сеголеток к старшим возрастным группам служит показателем тенденции изменения численности популяции. Это отношение определяется во время охотничьего сезона (осенью или зимой) по выборке из популяции, полученной охотниками или промысловиками. В общем, высокая численность молоди по сравнению с численностью взрослых указывает на то, что сезон размножения сложился успешно; и если смертность молоди не будет чрезмерной, то можно ожидать увеличения популяции в следующем году.
Рост популяции. Такие показатели, как рождаемость, смертность и возрастное распределение, весьма важны, но каждый их них сам по себе мало что дает; в частности, по каждому их них нельзя судить о росте популяции как целого, о характере изменений, которые произойдут при изменении условий среды, о возможностях популяции.
Внутренняя скорость роста.Если среда не налагает никаких ограничений (пространство, пища и другие организмы не оказывают лимитирующего действия), то скорость роста популяции для данных микроклиматических условий становится максимальной и постоянной. Значение скорости роста в этих условиях характеризуется соответствующей возрастной структурой и представляет собой единственный показатель наследственно обусловленной способности к росту - r.
dN/dt = rN, (1)
где: dN/dt- скорость изменения численности (скорость роста); r - скорость изменения численности в расчете на одну особь в единицу времени; N– численность.
Кривая роста с учетом действия лимитирующих факторов, как правило, имеет S - образный вид. Проинтегрировав уравнение (1) получаем экспоненциальную зависимость
Nt= N0 ert, (2)
где N0- численность в начальный момент времени, Nt - численность в момент времени t и e - основание натурального логарифма. После логарифмирования:
r = dN/(Ndt), (3)
r=(ln Nt – ln N 0)/t, (4)
если вместо Nt и N0 подставить Nt1 и Nt2 , вместо t - (t1 - t2) в фазе нелимитирующего роста получим скорость роста между двумя промежутками времени.

 b - const
b - const
 |
Удельная рождаемость (b)
 |
 Удельная смертность (d)
Удельная смертность (d)
 | |||
 |


Численность N пред
 b d
b d
b
 |
 d - const
d - const




Численность N пред
b d
b
 |

 d
d
Численность N пред
Рис. 3.8. Различные типы соотношений между удельной смертностью и рождаемостью
r = b - d (5)
Показатель r – фактически отражает разность между удельной скоростью рождаемости b(скорость рождаемости в единицу времени на одну особь) и удельной скоростью смертности – d. На рис.3.8. представлены различные типы соотношений между удельной смертностью и рождаемостью.
Общая скорость роста в нелимитирующей среде зависит от: возрастного состава, удельной скорости роста, удельной смертности.
Скорости роста для каждого вида могут быть различными в зависимости от структуры популяции. Максимальное значение r max называют биотическим или репродуктивным потенциалом.
 ЧИСЛЕННОСТЬ
ЧИСЛЕННОСТЬ
 |





 |
Дата добавления: 2014-02-26; просмотров: 754; Нарушение авторских прав

Мы поможем в написании ваших работ!