Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Зависимость видового насыщения от условий окружающей среды
Впервые зависимость видового насыщения определенна в фитоценотических правилах Жаккара и биоценотических принципах Тинеманна.
Жаккар объяснил видовое насыщение богатством и разнообразие растительного мира. По Жаккару видовое разнообразие зависит от ярусности леса, от широтной зональности и других факторов, объясняющих его разнообразие.
Правила Тинеманна связывают видовое богатство фитоценоза биоценоза с климатом.
Его положения:
1. видовое богатство достигается при максимальном разнообразии среды обитания. Разнообразие среды обусловлено определенными климатическими факторами.
2. Климат, в условиях которые достигаются разнообразие не должен быть суровым, хоть должен быть регулярно изменчивым. Видовой набор сообщества максимален на границе разных условий среды (опушка) и при условиях среды, при которых нет ее резких изменений.
В последствии были разработаны современные теории видового насыщения.
Гипотезы видового разнообразия
1. гипотеза опушки – на границе среды биоценозов наблюдается всплеск видового разнообразия относительно видовых наборов биоценозов (более разнообразные условия);
2. гипотеза хищничества – хищники способствуют видовому разнообразию (ослабляют межвидовую конкуренцию);
3. гипотеза сукцессивнного возраста экосистемы – количество видов экосистемы растет с увеличением ее возраста;
4. гипотеза экологических ниш, – чем больше локальных экологических ниш может образовать среда, тем больше видовой набор;
5. гипотеза конкуренции, – чем больше развит механизм внутривидовой конкуренции, тем она устойчивее;
6. гипотеза Мак-Артура, – чем богаче экосистема, тем она устойчивее.
Видовое разнообразие биоценоза является одним из главных факторов функционирования биогеоценоза или экосистемы, так как:
1. увеличивает число пищевых звеньев;
2. при видовом разнообразии растет поток энергии, проходящий через систему по разным пищевым цепям;
3. продуктивность зависит от видового разнообразия.
Чем разнообразнее условия существования в пределах биотопа, тем больше видов в данном биоценозе. И наоборот, чем больше отклонены от нормы условия существования в пределах биотопа, тем беднее видами становится биоценоз, тем больше особей одного вида. Таким образом, видовое разнообразие биоценоза достигается в условиях разнородной, но стабильной окружающей среды, рассмотренной как сумма биотической и абиотической компоненты.
ВЫВОД:
Видовое богатство биоценоза достигается при разнообразных условиях окружающей среды или богатстве и обилие фитоценоза, или при достаточно зрелых сообществах.
3.6.5 Видовое разнообразие
Сложность определения видового состава состоит в том, что группировки очень различаются между собой видовым разнообразием, которое является одним из основных показателей структуры биоценоза.
С одной стороны, охарактеризовать видовой состав группирования не сложно: перечислить виды, составить их список и, наконец, дать оценку видового богатство. С другой стороны, трудность состоит в том, что не все участки исследованного нами группирования имеют одинаковые показатели видового состава. Количество установленных видов будет зависеть от количества пробных площадей и их размеров.
Р.Уиттекер (1980) рассматривает несколько индексов видового разнообразия. Простейшим для определения является индекс разнообразия, где главным измерителем является количество видов S, описанных на площади стандартных размеров; S – это еще и единица измерения видового богатства, в отличие от плотности популяции.
Количество видов в выборках разного размера для данной группировки приблизительно пропорционально логарифму площади этих участков:
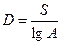 или
или 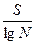 ;
;
где
D – это индекс разнообразия;
S – количество видов в описании на площадке стандартного размера;
A – площадь обчисляемой площадки;
N – общее количество особей в описании.
Лучшим для расчета является индекс разнообразия Симпсона, поскольку определяет для каждого вида часть его особей или биомассы в общем количестве. Если часть i-того вида Pi, то индекс разнообразия Симпсона:
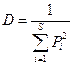 ;
;
где
S – общее количество видов в группировании (то есть видовое богатство).
Равномерность распределения (выравненность) можно так же количественно оценить с помощью индекса Симпсона, как часть максимально возможной величены D, которая достигается при одинаковом количестве всех видов.
Поскольку D max = S, то равномерность распределения:
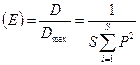
Этот показатель принимает значения от 0 до 1.
Индекс разнообразия Шеннона так же зависит от совокупности значений Pi:
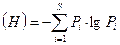
В этом случае равномерность распределения:
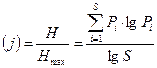
Оба показателя принимают максимальное значение, если части всех видов в группировании одинаковы.
Таким образом, с увеличением количества видов в группировке S максимальные значения обоих показателей, особенно показателя Шеннона, увеличиваются.
Поэтому для получения независимых от количества видов (S) характеристик обращаются к нормирования путем деления на максимальное значение того или иного показателя.
В связи с этим широко используют так называемый показатель выравненности:
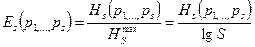
Этот показатель принимает значение от 0 до 1.
Видовое разнообразие группирований изменяется во времени и является следствием сложных процессов иммиграции и эмиграции видов, которые происходят в каждой экосистеме с большей или меньшей интенсивностью.
Новая наука палеоэкология дает возможность, изучая ископаемые останки животных и растительных организмов, установить каким был видовой состав группирований прошлого, проследить процесс их эволюции.
3.7 Трофическая структура сообщества
3.7.1 Трофические цепи и трофическая сеть сообщества
Последовательность питающихся друг другом организмов называется трофической цепью.
Особенности трофических отношений:
1. трофические цепи можно выделить искусственно для установления классификационных характеристик. В реальных условиях живые организмы переплетены в своих трофических отношениях, поэтому можно выделить только трофические уровни;
2. каждый трофический уровень составляют живые организмы, принадлежащие к разным видам, популяциям;
3. распределение живых организмов по трофическим уровням является функциональным и не связанным с их видовой принадлежностью;
4. многие организмы в дикой природе имеют смешанную диету (пример, лиса – виноград, олень – лемминги) и один и тот же организм в разных природных средах может занимать разные трофические уровни;
5. система трофических уровней с определенным видовым набором, составляющим каждый трофический уровень, составляет трофическую структуру сообщества.
3.7.2. Закон Линдеманна (правило 10%-ов)
Через трофические отношения в системе происходит перенос энергии, запасной через механизм фотосинтеза продуцентами всего сообщества.
В процессах поедания пищи происходит перенос энергии, заключенной в этой пище, по трофическим уровням. Этот процесс передачи энергии подчиняется законам термодинамики, то есть, каждый акт передачи энергии с уровня на уровень имеет (согласно II-ому закону термодинамики) свой КПД. Своеобразным выражением II-ого закона термодинамики в трофической системе является правило 10%-ов (закон Линдеманна):
При каждом акте переноса пищи через трофический уровень примерно 80 – 90% потенциальной энергии теряется, переходя в рассеянные формы и, таким образом, следующим трофическому уровню доступны лишь 10% энергии заключенной в пище.
Это правило обосновывает конечное число звеньев трофической цепи, так как вся энергия Солнца ассимилируется продуцентами и таким образом переданная для потребления в сообществе истощается примерно к четвертому уровню, поэтому наземные сообщества имеют 3-4 трофические уровни, водные – 5-6.
Основным параметром, от которого зависит количество энергии проходящей через трофический уровень, является эффективность потребления – это мера использования на одном трофическом уровне продукции предыдущего.
3.7.3. Пастбищные и детритные трофические цепи
Детритные цепи начинаются с мертвой органики (или подсистемы редуцентов).
В здоровом сообществе 90% всех трофических цепей являются детритными
Значения и функции детритных цепей:
1. детритные цепи превышают пастбищные по биомассе, численности организмов, составляющих трофические уровни, продуктивности и другим параметрам значимости. Например, существует общее правило: на суше биомасса грунтовых микроорганизмов и беспозвоночных всегда больше биомассы соответствующих организмов относящихся к пастбищным цепям;
2. подсистема редуцентов консервирует энергию для повторного использования;
3. такие функции других цепей определяют сложность и длину других пищевых цепей. Цепи редуцентов всегда длиннее и сложнее организованы;
4. пастбищные цепи неустойчивы сами по себе. Очень часто перевыпас растительноядных приводит к выеданию растительности сверх нормы и разрушению фитоценозов и всех пастбищных цепей, основаных на фитоценозах;
5. цепи редуцентов являются буфером природной системы. В них происходит увеличение накопительной буферной емкости экосистемы;
6. стабилизации биосферных циклов происходит через цепи редуцентов, являющихся обязательным элементом всех биологических круговоротов. Цепи редуцентов представляют собой пути возврата биогенов в окружающей природной среде.
3.7.4 Биологические пирамиды
Биологические пирамиды представляет собой график-гистограмму, которая отображает изменение определенных параметров по трофическим уровням сообщества.
Общую экологическую продуктивность сообщества и законы передачи энергии от уровня к уровню отражают 3 типа биологических пирамид (пирамиды Элтона):
1. продуктивности или энергии;
2. биомассы;
3. численности.
Биологическая пирамида – графическая модель, отражающая вещества и энергию, проходящую через сообщество.
Пирамиды продуктивности или энергии. Размерность ? или ? – для водных сообществ и летающих тварей. Идет учет вегетативных циклов растений. Пирамиды продуктивности всегда сужены к верху при условии, что все пищевые источники учтены, так как пирамиды являются отражением закона Линдемана.
Пирамиды биомассы. Биомасса может быть представлена в единицах веса и калорийности. Яркий пример перевернутой пирамиды (неправильного вида) – планктонные сообщества, в которых биомасса зоопланктона больше биомассы фитопланктона, но продуктивность фитопланктона настолько высока, что он может прокормить больше по массе зоопланктона. Нет мертвого планктона, это связано с особенностями разложения в данных сообществах. Пирамида биомассы отражает урожай на корню, а не скорость оборота биомассы. Кроме планктонных сообществ перевернутую пирамиду могут давать сообщества с быстрым размножением и высокой скоростью оборота, это дает возможность жертвам с низкой биомассой кормить более высокую биомассу при низкой скорости оборота.
Пирамиды численности (чисел). Отражают число особей на единицу площади или объема. Элтон считал, что одному хищнику необходимо для пропитания большее количество жертв. Так же как и пирамида биомассы она строится без учета времени на основе многих измерений. Пирамиды чисел отражают плотность особей и могут быть перевернутыми (групповая охота).
Закон пирамиды: из всех экологических пирамид только пирамида продуктивности зависит от скорости продуцирования пищи в противоположность пирамидам чисел и биомассы, отражающих статику системы, и характеризующим урожай на корню. Пирамида подробности отражает картину скоростей прохождения массы пищи через трофическую цепь, в соответствии с законом Линдемана эта пирамида всегда имеет правельную форму.
Ю.Одум в 1975 г. сформулировал закон пирамид: пирамида чисел приводит к переоценке значимости мелких особей сообщества, пирамида биомассы переоценивает роль крупных организмов, и лишь пирамида продуктивности отображают экологическую эффективность сообщества.
| <== предыдущая страница | | | следующая страница ==> |
| Индексы видового разнообразия | | | Общая характеристика и определение биосферы |
Дата добавления: 2014-08-09; просмотров: 559; Нарушение авторских прав

Мы поможем в написании ваших работ!