Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
МОДЕЛЬ ХОЛЛИДЕЯ
Рассмотрение гомологичной рекомбинации начнем с общей модели кроссинговера, опубликованной в 1964 году американским генетиком Р. Холлидеем. Холлидей предложил эту модель с конкретной целью для объяснения результатов, полученных им и другими исследователями генетики грибов. Она разработана для мейотического кроссинговера. Напомним, что ядро мейотической клетки в профазе I содержит по четыре гомологичных хроматиды, но в каждом отдельном акте кроссинговера участвуют только две из них. На рис. 58 изображены два гомологичных дуплекса, соответствующие хроматидам. Для простоты изображены только те две хроматиды из четырех, которые участвуют в кроссинговере.
В принципе для того, чтобы гомологичные молекулы ДНК поменялись своими частями, сначала должны произойти:
1. разрывы во всех цепях обоих дуплексов,
2. обмен цепями и замыкание разрывов. У Холлидея разрывы происходят не одновременно, а в два этапа:
а) Рекомбинация начинается с первичных одноцепочечных разрывов фосфодиэфирных связей ДНК (их вносит фермент эндонуклеаза). Разрывы происходят в двух цепях одинаковой полярности (рис. 58, а). Холлидей также постулировал, что первичные разрывы возникают не в случайных, а в определенных сайтах ДНК. Впоследствии эта идея получила экспериментальное подтверждение.
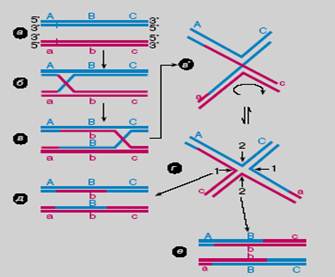
Рисунок 58. Модель Холлидея (Глазер В.М. 1998).
б) Далее от точек первичных разрывов происходит обмен цепями между дуплексами, который приводит к образованию крестообразной структуры, получившей впоследствии название "полухиазма Холлидея" (рис. 58, б и 59). Такое название объясняется тем, что в полухиазме в обмен вовлечены только две цепи ДНК из четырех, что отличает ее от полной хиазмы - характерного продукта завершенного мейотического кроссинговера, давно известного биологам.
3. Затем происходит очень важный процесс - перемещение точки перекреста цепей в полухиазме вдоль рекомбинирующих дуплексов (рис. 58, в). Такое явление описано под названием "миграция ветвления". Оно заключается в следующем:
а) от точки перекреста цепей происходит расплетание исходных дуплексов
б) высвобождающиеся цепи тут же ренатурируют с комплементарными цепями из гомологичных дуплексов,
в) что приводит к образованию и последующему удлинению гетеродуплекса (B / b на рис. 58, в). Именно в удлинении гетеродуплекса и заключается биологический смысл миграции ветвления. Ее осуществляют специальные ферменты. Размеры гетеродуплекса при мейотическом кроссинговере колеблются от нескольких сот до одной тысячи п.н., при рекомбинации в соматических клетках и клетках прокариот он еще протяженнее.
г) гетеродуплекс сформирован.
Образовавшаяся сложная разветвленная структура должна разделиться на гомологи. Это называется разрешением полухиазмы. Но прежде чем это случится, полухиазма должна претерпеть еще одно превращение - изомеризацию. Изомеризация заключается в изменении структуры полухиазмы, которое происходит за счет обычного теплового движения молекул.

Рисунок 59. Электронно-микроскопическая фотография - полухиазмы Холлидея (Глазер В.М. 1998).
На рисунке 58 изображена схема изомеризации полухиазмы, предложенная Х. Поттером и Д. Дресслером в 1976 году. Структуры в и в' идентичны: просто вторая изображена в крестообразном виде, что приближает ее к реальной (рис. 59). В структуре в' происходит один поворот на 180º любой пары дуплексных сегментов (плеч), на рисунке это нижняя пара. Образовавшаяся структура (рис. 58, г) может разрешиться двумя парами вторичных разрывов (вторичные разрывы завершат обмен цепями):
· Парные разрывы цепей одинаковой полярности 1-1 или 2-2 приводят к двум типам рекомбинантных хроматид: хроматиды первого типа (рис. 58, д ) содержат внутренний гетеродуплекс B / b, а по конфигурации фланговых маркеров А и С не отличаются от исходных (некроссоверные хроматиды);
· рекомбинантные хроматиды второго типа (рис. 58, е) кроссоверные, они также содержат гетеродуплекс, но обмениваются частями по обе стороны от него. Оба типа продуктов рекомбинации равновероятны, что соответствует генетическим данным, на которые опирался Холлидей при создании своей модели.
Здесь необходимо сделать небольшое отступление по поводу одного важного процесса, происходящего в гетеродуплексе. Как уже указывалось, от исходных молекул в рекомбинационный гетеродуплекс могут войти разные аллели, и тогда в нем возникнут неспаренные основания, которые локально нарушат структуру двойной спирали ДНК. Эти нарушения узнаются специальными ферментными системами, работающими по типу эксцизионной репарации. Они проводят коррекцию неспаренных оснований в гетеродуплексе: удаляют неспаренное основание в одной цепи ДНК и застраивают образующуюся брешь по матрице другого аллеля в комплементарной цепи, тем самым превращая (конвертируя) один аллель в другой. Это явление было давно известно под названием "конверсия гена", но теперь мы знаем, что в ее основе лежит коррекция гетеродуплекса.
Модель Холлидея в ее современном виде общепризнанна и универсальна для прокариот и эукариот (и для половых, и для соматических клеток). Ее достоинством является тот факт, что она хорошо проверяется генетическими данными, и практически все ее этапы постепенно нашли экспериментальное подтверждение. Полухиазмы Холлидея хорошо видны под электронным микроскопом (см. рис. 59). Обнаружены специальные эндонуклеазы (их называют резолвазами), которые осуществляют разрешение полухиазмы, как это изображено на рис. 3, г. К настоящему времени такие резолвазы обнаружены у бактериофагов T4 и T7, E. coli, дрожжей и человека. У E. coli выявлены также белки, осуществляющие миграцию ветвления полухиазмы. Именно на примере E. coli как наиболее изученного в отношении рекомбинации объекта мы рассмотрим конкретные механизмы кроссинговера.
| <== предыдущая страница | | | следующая страница ==> |
| Рекомбинантная система I - гомологичная генетическая рекомбинация | | | РЕКОМБИНАЦИЯ У E. COLI: ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ И МОЛЕКУЛЯРНЫЙ МЕХАНИЗМ |
Дата добавления: 2014-11-08; просмотров: 1151; Нарушение авторских прав

Мы поможем в написании ваших работ!