Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
питание и энергетический баланс животных
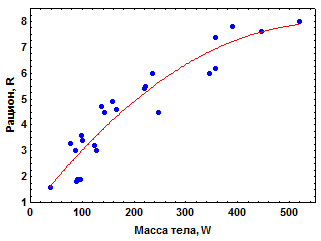
4.1. Зависимость рациона животных от массы их тела. Рацион любого организма (R), или количество пищи, потребленное за единицу времени, в значительной степени определяется массой его тела (W). Примером этого являются данные для кубинского краба Menippe mercenaria (Сущеня, 1975). Этот важный промысловый вид является падальщиком, который питается, преимущественно, остатками погибших животных.
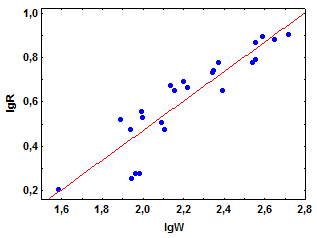
Если количество корма в среде находится с избытком, рацион крабов возрастает с увеличением массы их тела (рис. 4.1а). Однако увеличение рациона, как и скорости дыхания животных (см. гл. 3.2), происходит в значительно меньшей степени, чем массы тела. Поскольку в двойных логарифмических координатах (lgR - lgW) эмпирические значения рациона удовлетворительно располагаются вдоль прямой линии (рис. 4.1б), зависимость R от W у M. mercenaria может быть описана степенным уравнением
R = pWk , (4.1)
где p и k – эмпирические коэффициенты. При W = 1, р = R.
В численной форме уравнение (4.3) для M. мercenaria имеет вид:
R = 0,138W0.665, (4.2)
где R – рацион (г, сырая масса корма·особь-1сутки-1), W – сырая масса тела (г).
| 
|
Рис. 4.1. Зависимость среднесуточного рациона (R, г сырой массы корма особь-1) от сырой массы тела (W, г) у краба Menippe mercenaria (по Сущене, 1975)
Аналогичный характер связи R и W отмечен и для многих других видов ракообразных, моллюсков и представителей других таксонов. При этом значения степенного коэффициента k изменяются от 0,55 до 0,95, но обычно находятся в пределах 0,60 – 0,80. Например, обобщенное для класса ракообразных уравнение (4.1) имеет вид:
R = 0,0746W0,80,
где R – сырая масса корма·особь-1сутки-1, W – сырая масса тела, г (Сущеня, 1975).
Таким образом, характер зависимости среднесуточного рациона (т.е. скорости питания) у животных от массы их тела аналогичен таковому для скорости дыхания. Однако можно отметить, что степенные коэффициенты в уравнениях для скорости питания в целом несколько ниже, чем для скорости дыхания.
По уравнению типа (4.1) возможно рассчитать количество энергии, поступающее в организм в процессе питания, т.е. энергетический эквивалент потребленной пищи; однако для этого необходимо иметь данные об удельной энергоемкости вещества пищи.
Пример. У краба M.мercenaria массой 500 г среднесуточный рацион, рассчитанный согласно (4.2), составляет 8,6 г сырой массы. Соотношение между сухой и сырой массой его пищи равно 12%, а калорийность пищи – 4,5 ккал·г-1 сухого вещества. Отсюда краб с пищей за сутки потребляет в среднем 8,6 г·0,12·4,3 ккал·г-1 = 4,54 ккал энергии.
Если соотношение между сухой и сырой массой (γ) у пищевых объектов, потребляемых особями разных размеров и калорийность их сухой массы (Kγ) достаточно постоянны, уравнение (4.1) может быть представлено в виде:
R = γKγpWk.
Поскольку γKγp = q = const, полученное уравнение можно упростить:
R = qWk. (4.3)
Уравнение (4.3) представляет зависимость энергетического эквивалента потребленной организмом пищи от массы его тела.
4.2. Зависимость рациона от концентрации корма в среде. У всех видов животных, особенно у водных беспозвоночных с фильтрационным питанием (например, у жаброногих и ветвистоусых раков), величина рациона в значительной степени зависит от концентрации корма в среде. Как правило, при низких концентрациях корма рацион особей возрастает пропорционально их увеличению. Однако при дальнейшем повышении концентрации корма увеличение рациона все более замедляется, а выше определенного предела он стабилизируется на определенном уровне.
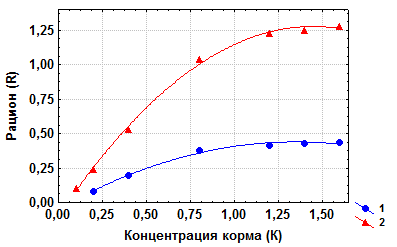
Для примера приведем данные по питанию жаброногого рака Artemia salina дрожжевыми клетками в условиях эксперимента (Сущеня, 1975). У обеих возрастных групп (молодь и взрослые особи) возрастание рациона с концентрацией корма происходил только до его величины, равной приблизительно 1,2 млн. клеток·мл-1, после чего уже практически не изменялся (рис. 4.2). У молоди максимальный рацион (Rmax) в среднем составлял 0,42 млн. клеток·особь-1час-1, а у взрослых особей – 1,24 млн. клеток·особь-1час-1.
|
Рис. 4.2. Зависимость между концентрацией дрожжевых клеток в воде (К, млн. клеток·мл-1) и рационом (R, млн. клеток-1·особь-1час-1) у Artemia salina. 1 – молодь; 2 – взрослые особи (по Сущене, 1975).
Полученная зависимость количества потребленного корма от его концентрации в среде может быть описана уравнением В.С.Ивлева (1955). Поскольку соотношение между реальным рационом (R) и концентрацией корма (К) пропорциональна разности между максимальным и реальным рационом (Rmax – R), получаем
dR/dK = λ(Rmax – R), (4.4)
где λ – коэффициент пропорциональности.
Интегрируя уравнение (4.4) , получаем асимптотическое уравнение
R = Rmax(1 – e-λK). (4.5)
В численной форме уравнение (4.5) для молоди A. salina имеет вид
R = 0,42 (1 – e-1,566K),
а для взрослых рачков:
R = 1,24(1 – e-1,267K),
где R – рацион (млн. клеток·особь-1час-1), К – концентрация корма (млн. клеток·мл-1).
По уравнению (4.5) возможно также рассчитать объем воды (V), очищаемый фильтраторами от водорослей. Для этого нужно величину рациона (R) особи разделить на соответствующую концентрацию водорослей в воде (K), т.е. V = R/K.
Пример. Одна взрослая артемия при концентрации корма в среде, равной 1 млн. клеток мл-1 потребляет 0,89 млн. клеток в час. Отсюда очищаемый ею объем воды равен 0,89 млн. клеток час-1 : 1 млн. клеток· мл-1 = 0,89 мл·час-1.
Кривая уравнения (4.5) проходит через начало координат, что предполагает, что животные способны потреблять корм даже при самом низком его содержании в среде. Это в целом справедливо для большинства растительноядных видов животных, а также хищников, падальщиков и детритофагов. Низкое количество корма в среде такие виды компенсируют увеличением затрат времени и энергии на его поиски, т.е., повышением активного обмена.
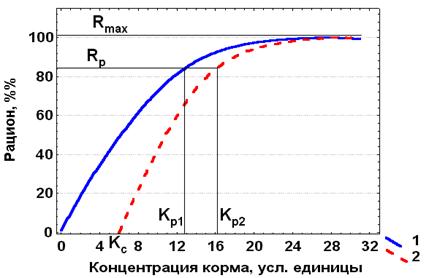
Однако многие виды с фильтрационным типом питания, например, ветвистоусые раки, если концентрации корма (планктонные одноклеточные водоросли) падает ниже определенного предела, вообще не способны его улавливать. Обычно чем грубее их фильтрационный аппарат, тем при более высоких концентрациях водорослей они теряют способность их потреблять.
Уравнение (4.5) для «грубых фильтраторов» может быть представлено в следующей форме, называемой уравнением Ивлева – Винберга – Анисимова
R = Rmax[1 – e-λ(K – Kс)].
Кривая зависимости рациона от концентрации корма у таких видов проходит не через начало координат, как на рис. 4.2, а через определенную точку на оси абсцисс (Кс > 0), соответствующую концентрации корма, выше которой животные становятся способными его улавливать (рис. 4.3).
4.3. Баланс энергии на организменном уровне. Экологические системы существуют за счет постоянного потока и трансформации вещества и энергии. Основными элементами экологических систем являются организмы. Поэтому особенности трансформации вещества и энергии в экологических системах в значительной степени определяются закономерностями этих процессов на организменном уровне.
Баланс энергии у гетеротрофных организмов можно схематично представить следующим образом. Потребленная пища, или рацион частично усваивается организмом (усвоенная, или ассимилированная часть рациона), а частично выделяется наружу в виде непереваренных остатков (неусвоенная часть рациона).
Усвоение пищи заключается в разложении ее сложных органических веществ (белков, углеводов, жиров) до сравнительно простых мономеров - аминокислот, моносахаридов и т.д. Одна их часть в процессе дыхания разлагается далее – до углекислого газа и воды с образованием АТФ. Процессы обмена веществ, ведущие к высвобождению энергии пищи в организме, имеют общее название энергетический обмен.
|
Рис. 4.3. Зависимость между концентрацией корма и рационом у видов с совершенным (1) и грубым (2) фильтрационным аппаратом (схема). Объяснение в тексте.
Другая часть мономеров используется для образования новых жиров, белков и углеводов, типичных для данного организма, которые затем используются на построение тканей и органов, т.е. на увеличение массы тела. Процессы образования в организме сложных органических веществ из более простых называется пластическим обменом.
Поскольку процессы пластического обмена могут происходить только с участием АТФ, оба типа обмена в организме тесно взаимосвязаны. Так, при синтезе белка в рибосомах на присоединении одной молекулы аминокислоты к строящейся полипептидной цепи затрачивается энергия одной макроэргической связи АТФ. Поскольку при окислении одной молекулы глюкозы образуется 38 молекул АТФ, этого количества достаточно для построения полипептидной цепи, состоящей из 39 аминокислот. Поэтому интенсивно растущие организмы потребляют много кислорода.
Согласно первому закону термодинамики, в любой системе между потреблением и расходом энергии существует равенство, т.е. соблюдается энергетический баланс. Для составления балансового равенства организма необходимо выразить каждый его компонент в единицах энергии. Баланс энергии организма можно представить в виде равенства:
R = P + T + N, (4.6)
где R – энергия потребленной пищи (рацион), Р – энергия пищи, использованная на увеличение массы тела особи (прирост), Т – энергия пищи, израсходованная в процессе дыхания, N – энергия пищи, которая потреблена организмом, но не усвоена им и выведена во внешнюю среду с фекалиями и другими продуктами распада.
Значения Т определяются по количеству кислорода, потребленного организмом, R – по энергетическому эквиваленту потребленной им пищи.
Сумма прироста и трат на дыхание в (4.6) является ассимилированной частью рациона (А)
А = Р + Т = RU-1,
где U-1 – усвояемость корма, или безразмерный коэффициент, представляющий собой отношение к рациону его усвоенной части, т. е.
U-1 = (P + T)/R = A/R.
По определению U-1 может изменяться в пределах от 0 до 1,00, а если ее выражают в процентах – от 0 до 100 %. У беспозвоночных и позвоночных животных, питающихся растительным кормом, U-1 , как правило, низка (до 20 – 50 %), что объясняется наличием в оболочках клеток растений трудно усваиваемой животными целлюлозы. У хищников, питающихся животной пищей, U-1 может быть значительно выше – до 80 %. У бактерий и грибов при потреблении отмершего органического вещества U-1 близки к 100 %, поскольку у них не образуются фекалии (Бигон и др., 1989).
В реальных условиях U-1, даже у отдельных видов, изменяются в очень широких пределах в зависимости от различных факторов – характера корма, температуры и других условий среды, физиологического состояния организма и т. д. Однако поскольку эти изменения не имеют, как правило, закономерного характера, для простоты дальнейших рассуждений U-1 для отдельных видов можно принять постоянной величиной.
Если усвоенную часть рациона организма представить в виде сумме А = Р + Т, то его рацион может быть определен согласно
R = (Р + Т)/U-1 . (4.7)
Приняв значение усвояемости пищи (U-1) постоянной величиной, из (4.1) получаем уравнение связи количества энергии, ассимилированной организмом (A), от массы его тела:
RU-1 = А = U-1qWk = q1Wk,
где q1 = U-1q.
Если в (4.7) принять Р = 0, то Т = RU-1 = A. Рацион организма, усвоенная часть которого равна его затратам энергии на дыхание, называется поддерживающим рационом (Rp). Очевидно,
Rp = T/U-1. (4.8)
Пример. Затраты особи на дыхание составляют 30 мгО2·час-1, усвояемость корма – 60 %, калорийность вещества корма – 1,2 кал·мг-1 сырого вещества. Отсюда поддерживающий рацион особи составляет 30 мгО2 час-1 · 3,39 кал мгО2-1 : 0,6 = 169,5 кал·час-1. Это эквивалентно 169,5 кал·час-1 : 1,2 кал·мг-1 сырого вещества = 141,3 мг сырого вещества·час-1.
При R = Rp масса тела организма остается постоянной, при R > Rp она будет возрастать, а при R < Rp – снижаться. Очевидно, длительное существование организма возможно лишь в случае, если его реальный рацион выше поддерживающего или хотя бы равен ему.
Пример. У краба M.мercenaria массой 500 г траты на дыхание, рассчитанные по уравнению для ракообразных, представленному в табл. 3.3, составляют 1,63 ккал· сутки-1. Усвояемость пищи у этого вида составляет 40%. Отсюда поддерживающий рацион составляет 1,63 : 0,40 = 4,08 ккал·сутки-1. Энергетический эквивалент пищи, потребленной крабом, рассчитанный выше, равен 4,54 ккал·сутки-1. Поскольку реальный рацион краба выше поддерживающего, масса его тела будет возрастать. Энергетический эквивалент суточного прироста массы тела равен 4,54 ккал – 4,08 ккал = 0,46 ккал·сутки-1.
У животных с фильтрационным типом питания концентрация корма, при которой рацион особей становится равным поддерживающему (Rp), называется поддерживающей концентрацией (Кс). Если два вида-фильтратора имеют одинаковый поддерживающий рацион, то поддерживающая концентрация корма (Кр) будет ниже для вида с более совершенным фильтрационным аппаратом (рис 4.3).
4.4. Экологическая эффективность роста. Любой гетеротрофный организм способен нормально существовать лишь тогда, когда поступление энергии с пищей будет превышать или хотя бы быть равным расходу энергии в процессе дыхания (то есть, когда А ≥ R). В этом случае будет происходить увеличение массы организма, т.е. его рост.
При А < R масса организма будет снижаться, поскольку вещества его тела расходуются в процессе дыхания.
Соотношение P/R = К1 называется валовой эффективностью роста, или коэффициентом эффективности роста первого порядка.
Соотношение P/А = К2 называется чистой эффективностью роста, или коэффициентом эффективности роста второго порядка.
Bсе параметры, используемые для расчетов K1 и K2, должны быть выражены в одинаковых единицах измерения, предпочтительнее – в энергетических единицах.
Согласно второму закону термодинамики, при любых формах перехода энергии определенная ее часть рассеивается в виде тепла. В организмах рассеяние энергии потребленной пищи происходит в процессах дыхания. Поэтому всегда К1 < 1 и K2 < 1. Поскольку А = RU-1 и К1 = К2/U-1, а U-1 < 1, то K2 > K1.
В практике рыбоводства и животноводства широко используется термин «кормовой коэффициент (КК)», или отношение прироста массы тела организма к массе потребленного им корма, т.е. КК = R/P = 1/K1. Иногда ихтиологи утверждают, что у рыб, питающихся комбикормом, КК может быть меньше единицы. Это означает, что прирост массы их тела может превышать массу съеденного ими корма, тогда К1 > 1,00.Однако это обусловлено тем, что прирост рыб выражен в сырой массе, а комбикорм – в сухой. При пересчете значений массы на их энергетические единицы оказывается, что KK всегда больше единицы.
Значения К2 у разных групп животных изменяется в очень широких пределах, однако в целом у пойкилотермных животных эти показатели значительно выше, чем у гомойтермных (табл. 4.1). Это обусловлено тем, что в энергетическом балансе млекопитающих и птиц доля затраты энергии на дыхание значительно выше, чем у пойкилотермных животных, что обусловлено значительно более высокого уровня метаболизма (табл. 3.4).
Таблица 4.1. Чистая экологическая эффективность роста (К2) у разных групп животных (по Humphreys, 1979 с изменениями)
| Группа организмов | К2 |
| Насекомоядные млекопитающие | 0,0086 |
| Другие млекопитающие | 0,015 – 0,031 |
| Птицы | 0,013 |
| Рыбы и общественные насекомые | 0,098 |
| Необщественные насекомые, эврифаги | 0,40 |
| -"-, фитофаги | 0,388 |
| -"-, детритофаги | 0,47 |
| -"-, хищники | 0,55 |
| Другие беспозвоночные, эврифаги | 0,25 |
| -"-, фитофаги | 0,208 |
| -"-, детритофаги | 0,276 |
| -"-, хищники | 0,362 |
У гомойотермных животных основным компонентом энергетического баланса являются затраты на дыхание, которые составляют до 90 % от величины ассимилированной энергии пищи. Поэтому значения К2 у них не превышают 1 – 3 %, а часто и гораздо меньше. Отсюда у растительноядных гомойотермных животных с низкими U-1 значения К1 не превышают 0,1 - 0,5%. У гомойотермных хищников К1 несколько выше – до 0,8 -1%.
У пойкилотермных животных затраты на дыхание существенно ниже. Поэтому значения К2 у них существенно выше и в среднем могут достигать 70 – 80%, а иногда и более. Поэтому энергия прироста массы их тела достигает 30% и более от энергии рациона.
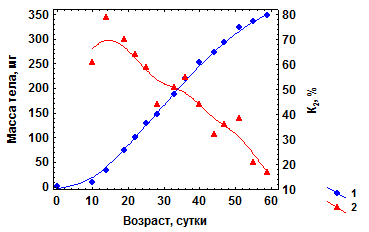
Значения К1 и К2 значительно изменяются в онтогенезе. Наивысшие значения К2 (до 0,85 – 0,9) отмечены на стадии эмбриогенеза. На постэмбриональных стадиях развития К2, как правило, снижаются с возрастом. Когда К2 становятся равными нулю, рост массы тела особей прекращается. На рис 4.4. представлены изменения К2 в течение всего жизненного цикла роста легочного моллюска физеллы Physella integra в водоеме-охладителе Березовской ГРЭС
|
Рис. 4.4. Изменения К2 в жизненном цикле у моллюска Physella integra, выращенного при 25 – 27 оС и оптимальной плотности (собственные данные). 1- средняя масса особей (W, мг); 2 – К2 , %
Видно что, несмотря на существенные колебания в процессе роста значения К2 в целом снижаются с возрастом, а у особей с предельной массой тела становятся близкими к нулю.
| <== предыдущая страница | | | следующая страница ==> |
| Тема 2.8.Электрический способ обогащения | | |
Дата добавления: 2014-11-14; просмотров: 628; Нарушение авторских прав

Мы поможем в написании ваших работ!