Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Тема 4. Строение, формы и свойства ДНК. Понятие гена, структура и свойства
А.1Высота капиллярного поднятия hс, м,–наибольшая (равновесная) высота подъема воды по порам грунта, отсчитываемая от зеркала грунтовых вод (равная мощности капиллярной каймы).
А.2Коэффициент водонасыщения Sr, д. е., определяется по формуле
| (А.1) |
где w — природная влажность грунта, д. е. (ГОСТ 5180);
е – коэффициент пористости, д.е.;
rs – плотность частиц грунта, г/см3 (ГОСТ 5180);
rw – плотность воды, принимаемая равной 1 г/см3.
А.3 Коэффициент выветрелости Кwr,д. е., определяется по формуле
| (А.2) |
где rВ - плотность выветрелого грунта, г/см3 (ГОСТ 5180);
rНВ - плотность невыветрелого грунта, г/см3 (ГОСТ 5180).
А.4 Коэффициент выветрелости крупнообломочных грунтов Кwrt, д. е., определяется по формуле
| (А.3) |
где К1 – отношение массы частиц размером менее 2 мм к массе частиц размером более 2 мм после испытания на истирание в полочном барабане;
К0 – то же, в природном состоянии.
А.5 Коэффициент истираемости крупнообломочных грунтов Кfr, д. е., определяется по формуле
| (А.4) |
где q1 – масса частиц размером менее 2 мм после испытания крупнообломочных фракций грунта (частицы размером более 2 мм) на истирание в полочном барабане;
q0 – начальная масса пробы крупнообломочных фракций (до испытания на истирание).
А.6 Коэффициент пористости е, д. е., определяется по формуле
| (А.5) |
где rs – плотность частиц грунта, г/см3 (ГОСТ 5180);
rd – плотность сухого грунта, г/см3.
А.7 Коэффициент размягчаемости в воде Кsof,д. е.,определяется по формуле
| (А.6) |
где Rс, Rвс – предел прочности грунта на одноосное сжатие соответственно в водонасыщенном и в воздушно-сухом состоянии (ГОСТ 12248).
А.8 Коэффициент сжимаемости мерзлого грунта mvf, МПа-1 – параметр, характеризующий объемную деформируемость мерзлого грунта под нагрузкой.
А.9 Коэффициент трещинной пустотности KТП,%-отношение суммарной площади трещин к площади породы.
А.10 Коэффициент фильтрации k, см/с или м/сут –скорость фильтрации воды при градиенте напора, равном единице, и линейном законе фильтрации(ГОСТ 25584).
А.11 Липкость (прилипаемость) – адгезионная прочность глинистых грунтов L,кПа – усилие, необходимое для отрыва плоского штампа из заданного материала от грунта после их контакта в течение заданного времени при определенном давлении.
А.12 Льдистость грунта за счет видимых ледяных включений ii,д. е.,определяется по формуле
| (А.7) |
где wtot – суммарная влажность мерзлого грунта, д. е. (ГОСТ 5180);
wm – влажность мерзлого грунта, расположенного между ледяными включениями, д. е.;
ww – влажность мерзлого грунта за счет содержащейся в нем при данной отрицательной температуре незамерзшей воды, д. е.;
ρs – плотность частиц грунта, г/см3 (ГОСТ 5180);
ρi – плотность льда, принимаемая равной 0,9 г/см3.
А.13 Относительная деформация набухания без нагрузки esw, д. е., – отношение увеличения высоты образца глинистого грунта при замачивании после свободного набухания в условиях невозможности бокового расширения к начальной высоте образца природной влажности (ГОСТ 12248).
А.14 Относительная деформация просадочности esl, д. е. – отношение разности высот образцов, соответственно, природной влажности и после его замачивания при заданном давлении (давление вышележащего грунта плюс давление от сооружения) к высоте образца природной влажности (ГОСТ 23161).
А.15 Относительное содержание органического вещества Ir, д. е. – отношение массы органического вещества к массе абсолютно сухого грунта (ГОСТ 23740 и ГОСТ 26213).
А.16 Плотность сухого грунта (скелета) ρd, г/см3, определяется по формуле
| (А.8) |
где r – плотность грунта, г/см3 (ГОСТ 5180);
w – естественная влажность грунта, % (ГОСТ 5180).
А.17 Показатель качества породы RQD,%, – отношение суммарной длины сохранных кусков керна длиной более 10 см к длине пробуренного интервала в скважине.
А.18Показатель текучести IL, д.е.– показатель состояния (консистенции) глинистых грунтов, определяется по формуле
| (А.9) |
где w – естественная влажность, % (ГОСТ 5180);
wP – влажность на границе раскатывания, % (ГОСТ 5180);
IP – число пластичности, %.
А.19 Показатель чувствительности грунта St,д.е., – отношение сопротивления недренированному сдвигу глинистых грунтов ненарушенного (сu) и нарушенного сложения (сur) или отношение сопротивления вращательному срезу (τmax) к его остаточному сопротивлению (τmin), определяется по формулам:
| или | (А.10) |
А.20 Пористость грунта n,%,определяется по формуле
| (А.11) |
где rs – плотность частиц грунта, г/см3 (ГОСТ 5180);
ρd – плотность сухого грунта, г/см3.
А.21 Предел прочности грунта на одноосное сжатие R,МПа– отношение нагрузки, при которой происходит разрушение образца, к площади его первоначального поперечного сечения (ГОСТ 12248).
А.22 Сопротивление недренированному сдвигу сu,кПа– величина, определяемая по результатам недренированных лабораторных и полевых испытаний глинистых грунтов (трехосные испытания, пенетрация, вращательный срез, зондирование и др.).
А.23 Степень засоленности грунта Dsal, % -отношение массы водорастворимых солей в грунте к массе абсолютно-сухого грунта.
А.24 Степень заполнения пор льдом и незамерзшей водой Sr, д. е., определяется по формуле
| (А.12) |
где wic – влажность мерзлого грунта, рассчитанная по содержанию порового льда, цементирующего минеральные частицы (лед-цемент), д. е., рассчитывается по формуле: wic=wm-ww;
ww – влажность мерзлого грунта, рассчитанная по содержанию незамерзшей воды при отрицательной температуре, д. е.;
rs – плотность частиц грунта, г/см3 (ГОСТ 5180);
еf – коэффициент пористости мерзлого грунта;
rw – плотность воды, принимаемая равной 1, г/см3.
А.25 Степень неоднородности гранулометрического состава Cu, д.е., определяется по формуле
| (А.13) |
где d60, d10 – диаметры частиц, мм, меньше которых в грунте содержится соответственно 60 и 10% (по массе) частиц.
А.26 Степень плотности песков ID, д.е.,определяется по формуле
| (А.14) |
где е – коэффициент пористости при искусственном сложении, д.е.;
emin – коэффициент пористости в предельно-плотном сложении, д.е. [5];
emax – коэффициент пористости в предельно-рыхлом сложении, д.е. [5].
А.27 Степень морозной пучинистости efh, % , определяется по формуле (ГОСТ 28622)
| (А.15) |
где h0,f – высота образца промерзшего грунта, см;
h0 – начальная высота образца грунта, см.
А.28 Степень разложения торфа Ddp,д. е., – отношение массы бесструктурной (полностью разложившейся) части к общей массе торфа (ГОСТ 10650).
А.29 Степень растворимости в воде qsr, г/л, – величина, отражающая способность грунта растворяться в воде при нормальных условиях за счет растворения неорганических и органических веществ, определяемая при соотношения грунта и воды 1:5 и равная концентрации образующегося равновесного раствора.
А.30 Суммарная льдистость мерзлого грунта itot, д. е.,определяется по формуле
| (А.16) |
где iic – льдистость грунта за счет льда-цемента (порового льда), д. е.;
wtot – суммарная влажность мерзлого грунта, д. е. (ГОСТ 5180);
ri – плотность льда, принимаемая равной 0,9 г/см3;
rs – плотность частиц грунта, г/см3 (ГОСТ 5180);
rf – плотность мерзлого грунта, г/см3 (ГОСТ 5180);
ww – влажность мерзлого грунта за счет незамерзшей воды, д. е.
А.31 Число пластичности Ip, % , определяется по формуле
| IP = wL – wP , | (А.17) |
где wL – влажность на границе текучести, % (ГОСТ 5180);
wP – влажность на границе раскатывания, % (ГОСТ 5180).
Тема 4. Строение, формы и свойства ДНК. Понятие гена, структура и свойства.
Развитие генетики на современном этапе связано с расшифровкой молекулярных основ наследственности. Все основные работы в этом направлении выполнены примерно за 20 лет. Расшифровка генетического кода позволила понять структуру и функции гена, а также изучить многие вопросы влияния гена на процессы онтогенеза. Вопросам реализации генетической информации у высших организмов при их развитии в онтогенезе уделяется особо большое внимание в современной молекулярной генетике. Все дело в том, что общие основы молекулярной генетики были заложены при изучении вирусов и бактерий, у которых генетическая структура, в сравнении с высшими организмами, устроена более просто.
ДНК была открыта Иоганном Фридрихом Мишером в 1869 году. Из остатков клеток, содержащихся в гное, он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
Первым, кто предположил, что генетическая информация заключена в хромосомах, был русский ученый Н. К. Кольцов (1927 г.). Однако он считал, что функцию хранителей наследственной информации выполняют специальные белковые молекулы, способные к автосинтезу. Кольцов был близок к решению проблемы генетического кодирования, но тщательные исследования показали, что информация заключена не на белковой молекуле, а на дезоксирибонуклеиновой кислоте (ДНК), которая представляет собой высокополимерное соединение, способное к автосинтезу.
Экспериментальные доказательства роли ДНК в передаче наследственной информации получили Освальд Эвери, Колин Мак-Леон и Маклин Мак-Карти (1944 г.). Они провели трансформацию, т.е. передачу наследственных признаков от одной группы бактерий к другой. В ходе эксперимента безвредная группа бактерий приобрела патогенные свойства, в результате добавления в неё мёртвых болезнетворных бактерий. Было показано, что за вновь приобретенные признаки отвечают выделенные из пневмококков ДНК.
Экспериментальные доказательства роли нуклеиновых кислот в передаче признаков позже получили американские учёные Алфред Херши и Марта Чейз на бактериофагах (эксперименты Херши-Чейз). Они вводили радиоактивные изотопы в белки и ДНК бактериофагов, и показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг.
Но точное строение ДНК, как и способ передачи наследственной информации, вплоть до 50-х годов XX века оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены.
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премии по медицине и физиологии в 1962 г. Среди лауреатов не было скончавшейся к тому времени Розалинды Франклин, так как премия не присуждается посмертно.
Дальнейшие работы были связаны с изучением принципа записи информации о строении белков на нуклеиновой кислоте, а также механизмов передачи этой информации к месту синтеза белка и следующему поколению при делении клетки. В 1961 году М. В. Ниренберг, Дж. Маттей , С. Очоа расшифровали и экспериментально подтвердили структуру генетического кода, закончив его исследование к 1964 году.
По современным представлениям, дезоксирибонуклеиновая кислота (ДНК) - макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК является своеобразной матрицей, на которой синтезируется информационная или матричная рибонуклеиновая кислота (м-РНК).
Локализация ДНК у организмов достаточно многообразна. У эукариотов ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах).
В клетках прокариотических организмов кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. И у прокариот и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
При характеристике величины ДНК организмов используются параметры генома. Геном – это общее содержание ДНК в гаплоидном ядре. Измерения размера генома приводятся в дальтонах, парах нуклеотидов (п. н.) или пикограммах (пг). Соотношение между этими единицами измерения следующие:
1 пг = 10–9 мг = 0,6х1012 дальтон = 0,9х109 п. н.
В гаплоидном геноме человека содержится около 3,2 млрд. п. н., что равно 3,5 пг ДНК. Таким образом, в ядре одной клетки человека содержится около 7 пг ДНК. Если учесть, что средний вес клетки человека равен примерно 1000 пг, то легко рассчитать, что ДНК составляет менее 1% от веса клетки. И, тем не менее, чтобы воспроизвести самым мелким шрифтом ту огромную информацию, которая содержится в молекулах ДНК одной нашей клетки, понадобилось бы тысяча книг по 1000 страниц в каждой. Размеры генома не говорят об уровне организации его обладателя. Например, у саламандры и лилии длина молекул ДНК, содержащихся в одной клетке, в тридцать раз больше, чем у человека.
Число отдельных молекул ДНК в клетке равно числу хромосом. Длина такой молекулы в наибольшей по размеру хромосоме 1 человека составляет около 8 см. Подобных гигантских полимеров пока не выявлено ни в природе, ни среди искусственно синтезированных химических соединений. У человека длина всех молекул ДНК, содержащихся во всех хромосомах одной клетке, составляет примерно 2 метра. Следовательно, длина молекул ДНК в миллиард раз больше их толщины. Так как организм взрослого человека состоит примерно из 5х1013 – 1014 клеток, то общая длина всех молекул ДНК в организме равна 1010 км (это почти в тысячу раз больше расстояния от Земли до Солнца).
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Другая известная нуклеиновая кислота – РНК, устроена аналогичным образом. Отличие состоит в том, что нуклеотиды РНК включают сахар рибозу.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). В РНК также встречается четыре вида азотистых оснований. Но вместо тимина присутствует урацил.
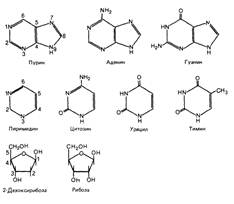
Азотистые основания представлены двумя группами: пиримидиновыми и пуриновыми. Основой пиримидинов является шестичленное пиримидиновое кольцо. Пурины имеют в своей структуре два слитых кольца- шестичленное и пятичленное.
Образование химических связей в нуклеотиде между его составляющими и отдельными нуклеотидами в полинуклеотидной цепи осуществляется через молекулу сахара. Поскольку в составе молекулы пентозы имеется 5 атомов углерода, то каждый из них можно пронумеровать индексом от 1' до 5'. В каждом нуклеотиде присоединение азотистого основания происходит к первому углеродному атому (1') пентозы с помощью β-гликозидной связи. Соединение, состоящее из углевода (пентозы) и азотистого основания, называется нуклеозидом (рис. ).
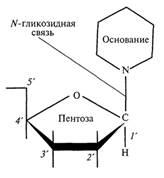
После присоединения к пятому углеродному атому остатка фосфорной кислоты образуется нуклеотид.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
Основание Нуклеозид Нуклеотиды (и принятые сокращения)
Аденин Аденозин Адениловая кислота = AMP (или dAMP)
Гуанин Гуанозин Гуаниловая кислота = GMP (или dGMP)
Цитозин Цитидин Цитидиловая кислота = СМР (или dCMP)
Тимин Тимидин Тимидиловая кислота = ТМР (или dTMР)
Урацил Уридин Уридиловая кислота = UMP (или dUMP)
ДНК имеет три уровня организации.
Первичная структура представлена полинуклеотидной цепью, скелетную основу которой составляют чередующиеся сахарные и фосфатные группы, связанные 3' - 5' – фосфодиэфирными ковалентными связями. Фосфоэфирные связи образуются между третьим и пятым атомами углерода соседних молекул дезоксирибозы в результате взаимодействия между 3'-гидроксильной (3'—ОН) группой одной молекулы дезоксирибозы и 5'-фосфатной группой (5'—РО3) другой. Этот остов имеет боковые группы, представленные азотистыми основаниями. Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3'-концу).
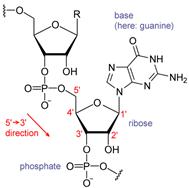
У большинства организмов (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК образует вторичную структуру, представленную двумя полинуклеотидными цепями, в которых азотистые основания ориентированы друг к другу. Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы. Эта двухцепочечная молекула спирализована. Именно, благодаря такому строению, молекула ДНК получила название «двойной спирали».
Связывание нуклеотидов двух нитей ДНК осуществляется за счет водородных связей. Азотистые основания одной из цепей соединены с азотистыми основаниями другой в строго определенном порядке согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями.
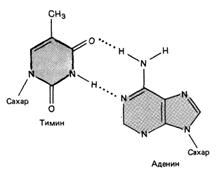

Последовательность пар нуклеотидов в ДНК нерегулярна, но сами пары уложены в молекуле как в кристалле. Это дало основание характеризовать молекулу ДНК как линейный апериодический кристалл.
При определении строения ДНК важную роль сыграло наблюдение, которое известно, как «правило Чаргаффа», согласно которым в каждой молекуле ДНК соблюдаются строгие соотношения, связывающие между собой количество азотистых оснований разных типов. Согласно правилам:
1.Сумма нуклеотидов, содержащих пуриновые основания, равна сумме нуклеотидов, содержащих пиримидиновые основания (А + Г = Т + Ц).
2.Независимо от действительного количества каждого основания относительное содержание Г всегда было равно относительному содержанию Ц, а содержание А - содержанию Т. Таким образом, любая ДНК может быть охарактеризована по ее составу как отношение (Г + Ц)/(А + Т), которое, варьируя от 26 до 74%, остается характерным для каждого вида.
Основные характеристики вторичной структуры ДНК:
1. Молекула ДНК образована двумя правозакрученными полинуклеотидными цепями, т.е. если смотреть вдоль оси спирали, повороты следуют по часовой стрелке. Ширина двойной спирали составляет от 22 до 24 Å, или 2,2 - 2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм), один поворот спирали состоит из 10 пар оснований. Каждая пара оснований повернута на 36º вокруг оси спирали относительно следующей пары оснований. Таким образом, 10 пар оснований составляют полный оборот в 360°. Две цепи, закручиваясь друг относительно друга, образуют двойную спираль, в которой имеется две бороздки - малая бороздка (около 12 Å шириной) и большая бороздка (около 22 Å шириной). Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны. Такая форма ДНК встречается в природе наиболее часто и она получила название В-форма.
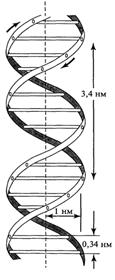
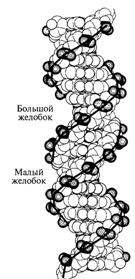
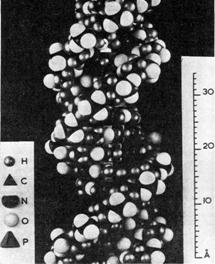
Рис. Модель вторичной структуры ДНК Уотсона Крика. Две цепи ДНК образуют двойную спираль.
2. Цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последовательности атомов в цепях обе цепи инвертированы относительно друг друга, т.е. направления от 3'-конца к 5'-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
5΄ фосфат 3΄ гидроксильная
группа
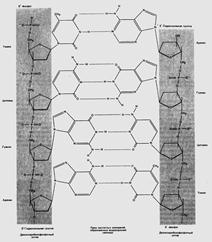
3΄ гидроксильная 5΄ фосфат
группа
Водородные связи между основаниями двух нитей могут разрываться. Разрыв водородных связей может происходить под воздействием различных факторов, что приводит к денатурации молекулы. Например, нагревание ДНК до 950 С вызывает разрушение водородных связей. Это явление называется плавлением ДНК.
Цепочки двойной спирали могут расходиться как замок-молния и под действием ферментов (хеликазы). Причем, на разрыв ГЦ требуется больше энергии, чем на разрыв пары АТ. Процент ГЦ пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки. Поэтому, части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, последовательности в бактериальных промоторах, обычно содержат большое количество А и Т.
Третичная структура ДНК определяется трехмерной пространственной организацией молекул и зависит от различных факторов.
| <== предыдущая страница | | | следующая страница ==> |
| Основные показатели строения, состава и свойств грунтов | | | Альтернативные структуры ДНК |
Дата добавления: 2014-04-16; просмотров: 744; Нарушение авторских прав

Мы поможем в написании ваших работ!