Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Редукционное деление
Профаза 1
Делится на ряд стадий, связанных с этапами прохождения кроссинговера.
Лептотена – стадия тонких нитей, связана с началом спирализации хромосом, которые становятся видны в виде клубка тонких нитей в диплоидном количестве. Каждая хромосома имеет вид тонкого, длинного волокна с белковой осевой нитью. В конце стадии в хромосомах становятся видны центромеры. На стадии лептотенны хромосомы прикреплены своими концами изнутри к СЯО особой структурой – прикрепительным диском. Хотя каждая хромосома уже реплицирована и состоит из двух сестринских хроматид, эти хроматиды так тесно сближены, что хромосома кажется одинарной (отдельные хроматиды не различимы вплоть до стадии диплотены или диакинеза).
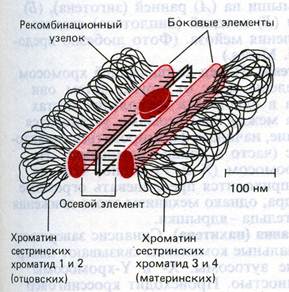
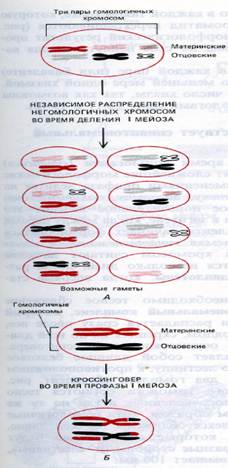
Зиготена – стадия конъюгации гомологичных хромосом, которые образуют биваленты. Момент перехода от лептотены в зиготену – начало синапса – тесной конъюгации гомологичных хромосом. Конъюгация начинается с того, что концы двух гомологичных хромосом сближаются на ядерной мембране, а затем процесс соединения гомологов продолжается вдоль хромосомы от обоих концов. При образовании бивалента каждый ген приходит в соприкосновение с гомологичным (аллельным) геном другой хромосомы. Когда гомологи конъюгируют, их белковые нити сближаются, образуя два боковых элемента длинного образования, напоминающего лестницу. Это образование называется синаптонемальный комплекс. Комплекс из двух сцепленных гомологичных хромосом называется бивалентом. Но правильнее эту структуру называть тетрадой, ведь в ее состав входит 4 сестринских хроматиды. В структуре бивалентов происходит исчезновение двойственной структуры пар гомологичных хромосом, поэтому количестов бивалентов – гаплоидное. В структуре бивалентов продолжается спирализация хромосом.
 Пахитена – стадия толстых нитей
Пахитена – стадия толстых нитей
Наступает после завершения процесса конъюгации гомологов. Пахитена может длиться несколько суток. Продолжение спирализации хромосом, в структурах бивалентов выявляются двухромосомная структура (4-х хроматидная - тетрады). На стадии пахитены происходит кроссинговер. В продольной щели синаптонемального комплекса появляются рекомбинационные узелки, которые вероятно играют большую роль в обмене участками между двумя несестринскими хроматидами между гомологами. Кроссинговер – обмен участками или фрагментами между гомологичными хромосомами. При наступлении кроссинговера происходит перекрест несестринских хроматид: в обмене участвуют по одной хроматиде из двух спаренных хромосом. В пахитене перекресты еще не видны, но позднее они проявляются в виде хиазм.
Диплотена – стадия двойных нитей
Начинается с расхождения, расталкивания и раскручивания гомологичных хромосом из состава бивалентов. Синаптонемальный комплекс распадается, что помогает двум гомологичным хромосомам бивалента несколько отодвинуться друг от друга.
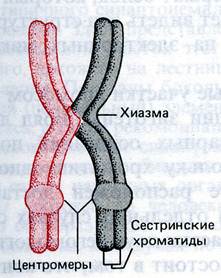 Однако гомологи остаются связанными друг с другом одной или несколькими хиазмами в точках прохождения кроссинговера. В бивалентах где произошел кроссинговер видны сползающие к концам хромосом хиазмы – перекресты между двумя хроматидами, претерпевшими кроссинговер.
Однако гомологи остаются связанными друг с другом одной или несколькими хиазмами в точках прохождения кроссинговера. В бивалентах где произошел кроссинговер видны сползающие к концам хромосом хиазмы – перекресты между двумя хроматидами, претерпевшими кроссинговер.
Одна из ососбенностей диплотенны – при некоторых типах оогенеза диплотенна может продолжаться очень долго (несколько лет). При этом, у некоторых организмов(например, у амфибий) происходит частичная деспирализация хромосом, возобновление транскрибционной активности (образование иРНК), так в этот период происходит цитоплазматический рост ооцитов. Такие хромосомы образуют многочисленные петли и называются хромосомы типа ламповых щеток.
Диакинез – завершение стадии профазы 1.
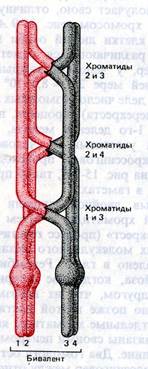 На этом этапе хромосомы – максимально спирализованы, синтез РНК прекращается. Хромосомы отделяются от ядерной оболочки. Ясно видно, что каждый бивалент состоит из 4-х отдельных хроматид. При этом каждая пара сестринских хроматид соединена центромерой, тогда как несестринские хроматиды, претерпевшие кроссинговер, соединены хиазмами. Разбирается СЯО, формируется веретено деления, биваленты перемещаются в область экваториальной пластинки.
На этом этапе хромосомы – максимально спирализованы, синтез РНК прекращается. Хромосомы отделяются от ядерной оболочки. Ясно видно, что каждый бивалент состоит из 4-х отдельных хроматид. При этом каждая пара сестринских хроматид соединена центромерой, тогда как несестринские хроматиды, претерпевшие кроссинговер, соединены хиазмами. Разбирается СЯО, формируется веретено деления, биваленты перемещаются в область экваториальной пластинки.
Остальные события мейоза занимают по времени не более 10% времени.
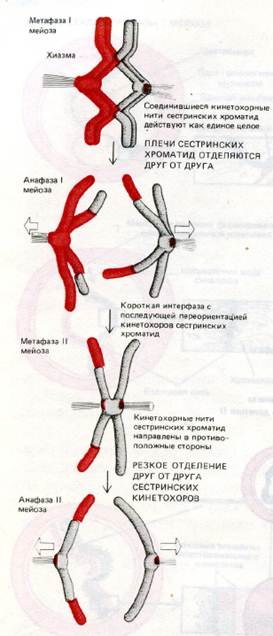
Метафаза 1.
В экваториальной области формируется метафазная пластинка из бивалентов хромосом.
Анафаза 1.
Расхождение целых гомологичных хромосом из состава бивалентов к полюсам, что приводит к редукции числа хромосом до гаплоидного. Расщепление хромосом из состава бивалентов происходит в плоскости конъюгации, поэтому в дочерние клетки попадают хромосомы отцовского и материнского происхождения, но случайно. Случайно означает, что к каждому полюсу отходит гаплоидный набор, но он может включать и отцовские и материнские хромосомы. Это второй уровень перемешивания генетического материала, следствием которого является то обстоятельство, что например дети похожи одновременно и на отца и на мать и одновременно на дедушку и на бабушку.
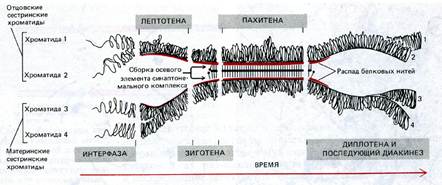
Схема, отражающая последовательность событий при конъюгации (синапсисе) и разъединении хромосом в профазе1 мейоза. Полностью сформированный синаптонемальный комплекс существует на протяжении всей стадии пахитены.
Телофаза 1.
Происходит по обычному сценарию и заканчивается образованием двух дочерних клеток с набором n2c.
Интерфаза – непродолжительна и в ней отсутствует фаза S (фаза редубликации ДНК).
Эквационное (выравнивающее) деление
Смысл этого деления – в завершении редукции количества ДНК до гаплоидного.
Второе деление происходит по схеме митоза, смысл этого деления в расхождении сестринских хроматид к полюсам.
 Профаза 1.
Профаза 1.
Короткая, так как хромосомы часто полностью не деспирализуются в интерфазе.
Метафаза 2.
В области экваториальной пластинки выстраиваются целые хромосомы в гаплоидном числе, состоящие из двух хроматид (после крссинговера).
Анафаза 2.
К полюсам клетки расходятся половинки хромосом (хроматиды) неидентичные друг другу.
Телофаза 2.
Завершается образованием полностью гаплоидных клеток и по числу хромосм и по количеству ДНК.
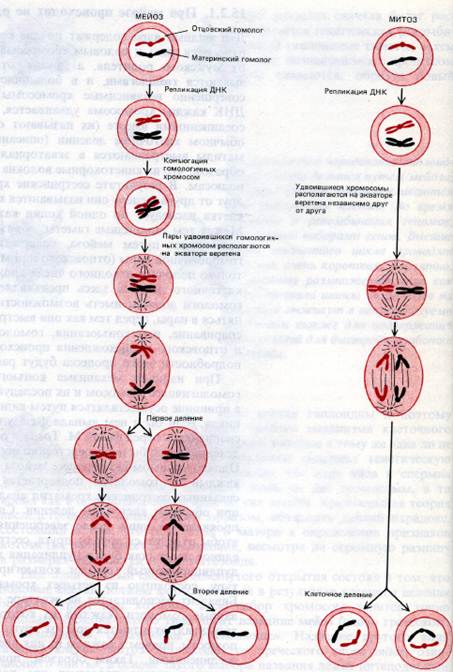
Схема на которой сравниваются механизмы упорядоченного расположения хромосом в метафазе и механизмы расхождения хромосом в первом и втором делении мейоза. Во втором делении мейоза используются те же механизмы, что и при обычном митозе.
Ниже показаны схемы мейоза и сравнения мейоза с обычным митотическим делением.
Для простоты в обеих делениях показана только одна пара гомологичных хромосом.
Спаривание (конъюгация) гомологичных хромосом и кроссинговер происходят только в мейозе. Мейоз в отличие от митоза совершается за два клеточных деления, результатом которых является образование 4-х гаплоидных клеток.
| <== предыдущая страница | | | следующая страница ==> |
| История открытия мейоза | | | Цитология. Энергетический обмен веществ |
Дата добавления: 2014-09-10; просмотров: 1363; Нарушение авторских прав

Мы поможем в написании ваших работ!