Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
ТЕМА: МЫШЕЧНЫЕ ТКАНИ
Цели занятия. Научиться:
1.Определять на светооптическом уровне исчерченную, неисчер-ченную(гладкую),сердечную мышечные ткани.
2.Узнавать и анализировать отличия разных видов мышечной ткани.
3.Анализировать на электронно-оптическом уровне структуру мышечного волокна, гладкой мышечной клетки, кардиомиоцитов.
Методические рекомендации к изучению темы: повторить общие вопросы раздела «Миология» из анатомии человека и ответить на следующие вопросы:
1.Что является источником формирования исчерченной мускулатуры органов опоры и движения?
2. Каково функциональное назначение скелетных мышц?
3. Принципы работы мышц.
4.Строение скелетной мышцы как органа.
5.Из каких мышц построены стенки полых внутренних органов?
Имея разное происхождение и строение, мышечные ткани объединяет способность к сокращению. Сократительный аппарат занимает значительную часть в цитоплазме, в его составе присутствуют актиновые и миозиновые филаменты, которые формируют органеллы специального значения–миофибриллы.
По морфофункциональному признаку различают:
· Скелетную или поперечно-полосатую или исчерченную мышечную ткань. Начало и прикрепление мышц находится на скелете. Мышцы являются произвольными, поскольку их сокращения подчиняются нашей воле. К этой группе мышц относят скелетные мышцы, мышцы языка, гортани и др.
· Сердечная мышечная ткань входит в состав мышечной стенки сердца. Иннервируется вегетативными нервами, является непроизвольной.
· Гладкая (неисчерченная) мышечная ткань характеризуется отсутствием исчерченности, а поскольку сокращения также не подчиняются нашей воле, то мышцы называют непроизвольными. Иннервация осуществляется вегетативной нервной системой. Из гладких мышц построены стенки внутренних органов, стенка сосудов.
В зависимости от источников развития выделяют пять типов мышечной ткани:
1. Мезенхимного происхождения (гладкая мышечная ткань).
2.Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки (например, в потовых, слюнных железах).
3.Нейральное происхождение (из нервной трубки)–мышцы суживающие и расширяющие зрачок.
4.Целомическое происхождение (миоэпикардиальная пластинка) – сердечная мышечная ткань.
5. Из миотомов мезодермы– исчерченная мышечная ткань.
Исчерченная (поперечно-полосатая) мышечная ткань Источником развития являются клетки миотомов миобласты. Различают головные, шейные, грудные, поясничные, крестцовые миотомы. Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов.
Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие, с 3 недели внутриутробного развития мигрируют в мезенхиму и сливаясь, друг с другом образуют мышечные трубки (миотубы) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают. Гранулярная ЭПС значительно редуцируется. Такая многоядерная структура называется симпласт, а для мышечной ткани - миосимпласт.
Часть миобластов дифференцируется в миосателлитоциты, которые располагаются на поверхности миосимпластов и принимают участие в регенерации мышечной ткани.
Структурной единицей мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной (рис.21). Длина мышечных волокон колеблется от 1 до 40 мм, а толщина 0,1 мм.
В мышечном волокне различают мембранный аппарат, фибриллярный (сократительный) аппарат, трофический аппарат (ядро, саркоплазма, цитоплазматические органеллы).
Мембранный аппарат. Каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), плазмолемма образует впячивания (Т- трубочки).
К плазмолемме снаружи прилежат миосателлитоциты. При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов.
Фибриллярный аппарат. Исчерченные волокна можно разделить на составляющие их фибриллы (диаметром 1 мкм), названные миофибриллами. В мышечном волокне они ориентированы продольно.
При рассматривании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками или А- дисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными или I – дисками. В средней части диска А имеется более светлый участок Н-зона (участок содержащий только толстые нити белка миозина). В области Н-зоны выделяется более темная М-линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z, которая построена из белковых фибриллярных молекул. В частности, большую роль играет альфа актинин. Z – линия соединена с соседними миофибриллами с помощью белка десмина и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечно-полосатой исчерченности мышечного волокна.
Структурной единицей миофибриллы является саркомер (S) - это пучок миофиламентов заключенный между двумя Z линиями (рис.22). Принимая во внимание вышеуказанные обозначения можно структуру саркомера записать в виде формулы:
S= Z1 + 1/2 I1 + А + 1/2 I2 + Z2
Под электронным микроскопом миофибриллы представляют агрегаты из толстых (меозиновых) филаментов ( диаметр 14 нм, длина 1500 нм, расстояние между ними 20-30 нм). Между толстыми филаментами располагаются тонкие филаменты ( диаметр 7-8 нм).
Толстые филаменты (миозиновые нити) состоят из молекул белка миозина. Он является важнейшим сократительным белком мышцы. При непосредственном участии миозина химическая энергия трансформируется в механическую работу. Каждая миозиновая нить состоит из 300-400 молекул миозина. Молекула миозина – это гексамер, состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах глобулярные (шаровидные) головки. Между головкой и тяжелой цепью – шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок - легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити.
Тяжелые и легкие цепи в молекуле миозина можно разделить обработкой мочевиной, гуанидинхлоридом и др. При мягкой обработке можно отделить только легкие цепи. Миозину свойственна АТФ-азная активность – высвобождающаяся энергия используется для мышечного сокращения.
Тонкие нити (актиновые нити). Образованы тремя белками: актином, тропонином и тропомиозином. Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали.
Толстые нити занимают центральную часть саркомера–А-диск, тонкие занимают I диски и частично входят между толстыми миофиламентами. Только толстые нити содержит Н-зона.
При поступлении нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна. Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров, располагающихся в прилежащей рыхлой соединительной ткани. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном называется нервно-мышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон).
В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие участки актина заблокированы тропомиозином. При высокой концентрации ионов кальция конформационные (пространственные) изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина. Плазмолемма миосимпласта образует пальцевидные впячивания (инвагинации) ориентированные поперечно по отношению к миосимпласту называемые Т-трубочки. К каждой Т-трубочке примыкают по две цистерны саркоплазматического ретикулума (гладкая ЭПС), образуя триаду: две цистерны и одна Т-трубочка. Са2+ концентрируется в цистернах ( там его концентрация в 800 раз больше, чем в саркоплазме).
Механизм сокращения. При поступлении нервного импульса волна деполяризации доходит до цистерн саркоплазматического ретикулума, из них выделяются ионы кальция и концентрация кальция в саркоплазме резко возрастает. Са2+ диффундирует к тонким нитям (филаментам) саркомера, где связывается с тропонином и миозиновыми головками. Это приводит:
1.К изменению конформации (пространственного расположения) тропомиозина, что, в свою очередь, приводит к освобождению участков актина, необходимых для взаимодействия с миозиновыми головками.
2.Появлению АТФ-азной активности миозина.
3.Взаимодействию миозиновых головок с актином (актино- миозиновые «мостики»).
Все это вместе взятое приводит к тому, что миозиновые головки «шагают» по актину, образуя в ходе перемещения новые связи актина и миозина, сближая две Z-линии. При сокращении уменьшаются только светлые диски.
Расслабление. Са2+-АТФ-аза саркоплазматического ретикулума закачивает Са2+ из саркоплазмы в цистерны. В саркоплазме концентрация Са2+ становится низкой. Са2+-тропомиозин закрывает миозин-связывающие участки тонких нитей и препятствует их взаимодействию с миозином.
Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон изменяется натяжение соединительно-тканной капсулы веретена и соответственно изменяется тонус интрафузальных (расположенных под капсулой) мышечных волокон. Формируется нервный импульс.
Классификация и типы мышечных волокон. Скелетные мышцы, состоящие из мышечных волокон отличаются по многим параметрам: цвету, диаметру, утомляемости, скорости сокращения и т.д. В каждой мышце присутствуют разные типы мышечных волокон. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные, которые преобладают и обуславливают собственно сократительную функцию мышцы и интрафузальные, входящие в состав проприоцепторов–нервно-мышечных веретен.
По характеру сокращения мышечные волокона делят на фазные и тонические. Фазные способны осуществлять быстрые сокращения, но не могут длительно удерживать достигнутый уровень укорочения. Тонические –обеспечивают поддержание статического напряжения или тонуса.
По биохимическим особенностям и цвету выделяют красные и белые мышечные волокна. Цвет мышцы обусловлен степенью васкуляризации. Кроме того, существует прямая корреляция между содержанием миоглобина и цветом мышцы. Характерной особенностью красных мышечных волокон является наличие многочисленных митохондрий, цепи которых располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше и они располагаются равномерно в саркоплазме мышечного волокна.
Скорость сокращения определяется типом миозина. Высокую скорость сокращения обеспечивает быстрый миозин (для него характерна высокая активность АТФ-азы); меньшая скорость сокращения характерна для медленного миозина (характерна невысокая активность АТФ-азы). Следовательно, по активности АТФ-азы можно судить и о наборе миозинов.
Тип окислительного обмена. Мышечные волокна используют два пути образования АТФ:
*при анаэробном типе метаболизма из 1 молекулы глюкозы образуется 2 молекулы АТФ и молочная кислота.
*при аэробном окислении из 1 молекулы глюкозы образуется 38 молекул АТФ и конечные продукты метаболизма: СО2 и Н2О. Идентификация мышечных волокон основана на выявлении активности фермента сукцинатдегидрогеназы (СДГ), которая является маркером для митохондрий и цикла Кребса. Активность этого фермента свидетельствует о напряженности энергетического метаболизма. Выделяют мышечные волокна А-типа (гликолитические) с низкой активностью СДГ, С-тип (оксидативные) с высокой активностью СДГ. Мышечные волокна В-типа занимают промежуточное положение. Переход мышечных волоко от А-типа в С-тип маркирует изменения от анаеробного гликолиза к метаболизму, зависящему от кислорода.
Существует много и других классификаций.
Факторами, определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное влияние, уровень васкуляризации, уровень двигательной активности и местоположение мышцы.
Сердечная мышечная ткань находится в мышечной оболочке сердца (миокард) и в устьях связанных с ним крупных сосудов. Имеет клеточный тип строения и основным функциональным свойством служит способность к спонтанным ритмическим сокращениям.
Развивается из миоэпикардиальной пластинки (висцеральный листок спланхнотома в шейном отделе), клетки которой размножаются митозом а потом дифференцируются. В клетках появляются миофиламенты, которые далее формируют миофибриллы.
Сердечная мышечная ткань образована клетками, которые называются кардиомиоциты. Между ними расположена рыхлая соединительная ткань и кровеносные сосуды с помощью вставочных дисков. Кардиомиоциты обьединяются в мышечные «волокна». Продольные и боковые связи кардиомиоцитов обеспечивают функциональное единство миокарда. Последние являются комплексом контактов. На поперечном срезе вставочных дисков выявляют десмосомы и щелевидные контакты (нексусы).
Выделяют рабочие (сократительные) кардиомиоциты, которые образуют цепочки клеток и обеспечивают силу сокращения сердечной мышцы. Клетки удлиненной формы с центрально расположенным ядром (рис.23). Вблизи ядра (или двух) комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Имеются Т-трубочки и L-трубочки. Десмосомы обеспечивают механическое сцепление, которое препятствует расхождению кардиомиоцитов. Щелевидные контакты способствует передаче сокращения от одного кардиомиоцита к другому.
Проводящие (атипичные) кардиомиоциты–среди них различают: 1.Водители ритма–это клетки небольших размеров, в саркоплазме мало гликогена, мало миофибрилл и они расположены по периферии. Клетки имеют хорошее кровоснабжение и иннервацию. Они воспринимают сигналы от нервных окончаний и способны автоматически генерировать сигналы обеспечивающие ритмические сокращения сердца.
2.Проводящие (переходные) кардиомиоциты проводят возбуждение от водителя ритма. Образуют длинные волокна. Миофибриллы в небольшом количестве, имеют спиральный ход, мелкие митохондрии, немного гликогена.
3.Волокна Пуркинье–являются самыми крупными клетками в мышечной ткани сердца с неупорядоченным расположением миофибрилл, множеством мелких митохондрий, много гликогена, нет Т-трубочек, клетки связаны между собой десмосомами и щелевидными контактами.
Секреторные кардиомиоциты – находятся в, основном, в предсердиях, преимущественно в правом. Характеризуются отростчатой формой и слабым развитием сократительного аппарата. В саркоплазме, вблизи полюсов ядра-секреторные гранулы, содержащие атриопептин (гормон, регулирующий артериальное давление). Гормон вызывает потерю натрия и воды с мочой, расширение сосудов, снижение давления, угнетение секреции альдостерона, кортизола, вазопрессина.
Сократительный аппарат рабочих кардиомиоцитов сходен со скелетными мышечными волокнами. Миофибриллы в кардиомиоците могут объединяться в комплексы, образуя единые сократительные структуры. В саркоплазме миофибриллы ориентированы продольно и располагаются преимущественно по периферии. Саркотубулярная система вцелом имеет сходство с исчерченными мышечными волокнами. Саркоплазматическая сеть развита слабее, не так активно накапливает Са2.. При расслаблении, ионы кальция выделяются в саркоплазму с низкой скоростью, что обеспечивает автоматизм и частые сокращения кардиомиоцитов. Т-трубочки широкие и образуют диады (одна Т-трубочка и одна цистерна сети), которые сходятся в области Z-линии. Энергетический аппарат-это митохондрии и включения.
Неисчерченная (гладкая) мышечная ткань. Структурно-функциональной единицей данной ткани являются гладкие мышечные клетки ( ГМК), которые способны к гипертрофии и регенерации. Они образуют стенки внутренних полых органов, сосудов. Более крупные по размерам ГМК характерны для стенок полых внутренних органов, а меньших размеров–для стенки сосудов. Клетки контролируют моторику, величину просвета. Имеют веретенообразную форму, в центре палочковидной формы ядро. В ГМК отсутствует поперечно-полосатая исчерченность. ГМК окружены сарколеммой, которая снаружи покрыта базальной мембраной. Длина от 20 мкм до 1 мм. В саркоплазме у полюсов находится комплекс Гольджи, много митохондрий, рибосом, развит саркоплазматический ретикулум. Миофиламенты расположены вдоль продольной оси. В ГМК актиновые и миозиновые филаменты не формируют миофибрилл. Актиновые нити (тонкие филаменты) ориентированы по продольной оси ГМК. По количеству их больше и они прикрепляются к плотным тельцам, которые являются специальными сшивающими белками. Рядом с актиновыми нитями располагаются мономеры миозина (микромиозин). Обладая разной длиной они, значительно короче тонких нитей.
Сокращение гладких мышечных клеток осуществляется при взаимодействии актиновых филаментов и миозина. Сигнал идущий по нервным волокнам обуславливает выделение медиатора, что изменяет состояние сарколеммы. Она образует колбовидные впячивания (кавеолы), где концентрируются ионы кальция. Сокращение ГМК индуцируется притоком ионов кальция в саркоплазму (кавеолы отшнуровываются и вместе с ионами кальция попадают в саркоплазму). Это приводит к полимеризации миозина и взаимодействию его с актином. Актиновые нити и плотные тельца сближаются, усилие передается на сарколемму и ГМК укорачивается. Миозин ГМК способен взаимодействовать с актином только после фосфорилирования его легких цепей особым ферментом–киназой легких цепей. После прекращения сигнала ионы кальция покидают кавеолы; миозин деполяризуется, теряет сродство к актину. В результате комплексы миофиламентов распадаются; сокращение прекращается.
Особые типы гладких мышечных клеток. Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности. Окружают секреторные отделы и выводные протоки желез (слюнных, молочных, слезных). С железистыми клетками они связаны десмосомами. Сокращаясь, способствуют выделению секрета. В концевых (секреторных) отделах форма клеток отросчатая, звездчатая. Ядро в центре, в цитоплазме, преимущественно в отростках локализованы миофиламенты, которые образуют сократительный аппарат. В этих клетках есть и цитокератиновые промежуточные филаменты, что подчеркивает их сходство с эпителиоцитами.
Мионейральные клетки–развиваются из клеток наружного слоя глазного бокала и образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок. По строению первая мышца сходна с ГМК мезенхимного происхождения. Мышца, расширяющая зрачок образована отростками клеток, располагающимися радиально, а ядросодержащая часть клетки находится между пигментным эпителием и стромой радужки.
Миофибробласты относятся к рыхлой соединительной ткани и представляют собой видоизмененные фибробласты. Они проявляют свойства как фибробластов, так и ГМК (обладают выраженными сократительными свойствами). Как вариант этих клеток можно рассматривать миоидные клетки в составе стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника. При заживлении раны часть фибробластов синтезирует гладкомышечные актины и миозины.
Эндокринные гладкие миоциты–это видоизмененные ГМК, представляющие основной компонент юкста-гломерулярного аппарата почек. Они находятся в стенке артериол почечного тельца, имеют хорошо развитый синтетический аппарат и редуцированный сократительный. Продуцируют фермент ренин, находящийся в гранулах и попадающий в кровь механизмом экзоцитоза.
Вопросы для самоконтроля:
1.Как классифицируются мышечные ткани по морфо-функциональному признаку? по происхождению?
2.Что является структурно-функциональной единицей мышечной ткани?
3.Строение фибриллярного аппарата мышечных волокон.
4.Напишите формулу саркомера.
5.Строение мышечных волокон под световым и электронным микроскопом.
6.Механизм сокращения и расслабления мышечного волокна.
7.Как классифицируются мышечные волокна? Типы мышечных волокон?
8.Виды кардиомиоцитов в сердечной мышце, особенности их строения.
9.Строение ГМК.
10.Перечислить особые типы гладких мышечных клеток.
Ситуационные задачи.
1. При обработке экспериментального материала (одна из скелетных мышц крысы) выявлено по активности СДГ три типа мышечных волокон: А, В, С. Какой тип метаболизма характерен для каждого из этих мышечных волокон?
2. Какие наследственные и ненаследственные заболевания мышц Вы знаете?
3. Как бы Вы обьяснили механизмы, происходящие при нарушении питания участка сердечной мышцы?
4.В судебном морге на медицинскую экспертизу представлены препараты исчерченной мышечной ткани. В одном препарате выявлены симпласты с ядрами расположенными по периферии, а в другом–клетки с центрально ориентированными ядрами. Каким органам принадлежат выявленные мышечные ткани?
5.У больного инфаркт миокарда, повреждена сердечная мышца. Какие клетки обеспечат восстановление дефекта в сердечной мышце?
6. ТЕМА: НЕРВНАЯ ТКАНЬ
Цели занятия:
1. Обобщить изученный материал, выделяя наиболее важные аспекты темы.
2. Научиться узнавать на светооптическом уровне структуры нервной ткани (нейроны, нервные волокна, нейроглию).Анализировать их строение.
3. Анализировать на электронно-оптическом уровне структуру нейронов, синапсов, нервных волокон.
Повторите вопросы из предшествующих тем:
1. Цитоскелет клетки. Значение микротрубочек в транспортных процессах в клетке.
2. Рецепторы клетки.
3. Облегченная диффузия и активный транспорт. Калий-натриевый насос.
4. Механизмы экзоцитоза.
5. Межклеточные контакты.
6. Строение и значение митохондрий, гранулярной эндоплазматической сети, пластинчатого комплекса.
7. Понятие о стабильной клеточной популяции.
8. Механизмы миграции клеток.
9. Макрофаги и макрофагическая система.
Общая характеристика нервной ткани. Нервная ткань (рис.24) выполняет функции восприятия, проведения и передачи воздуждения, полученнного из внешней среды и внутренних органов, а также анализ, сохранение полученной информации, интеграцию органов и систем, взаимодействие организма с внешней средой. Развивается из дорзальной эктодермы на первом месяце внутриутробной жизни.
Основными структурными элементами нервной ткани являются нейроны с их отростками и нейроглия. Нейроглия в свою очередь делится на микроглию и макроглию. Макроглия центральной нервной системы подразделяется на следующие клетки: астроциты (волокнистые и протоплазматические), олигодендроциты и эпендимоциты. Макроглия периферической нервной системы включает в себя сателлитоциты и леммоциты.
Нейроны, выполняют основные функции, приписываемые нервной ткани. Они состоят из тела (перикариона) и отростков. При этом последние могут многократно превышать объем тела нервной клетки. Среди отростков выделяют дендриты и аксон (нейрит). Дендриты сильно ветвятся, образуя дендритное дерево, и обычно короче аксона. От дендритов возбуждение направляется к телу нервной клетки. Они формируют постсинаптические структуры, воспринимающие возбуждение. Дендритов много, но может быть один. Аксон присутствует всегда, по одному на каждую нервную клетку. Он не ветвится или слабо ветвится в терминальных областях и заканчивается синаптическим бутоном, передающим возбуждение на другие клетки (пресинаптическая зона). Нейроны передают возбуждение с помощью специализированных контактов (синапсов). Вещество, обеспечивающее передачу возбуждения, называется медиатором. В каждом нейроне обычно обнаруживается один основной медиатор. Нейроны являются морфологической и функциональной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами, образуя рефлекторные дуги.
 |
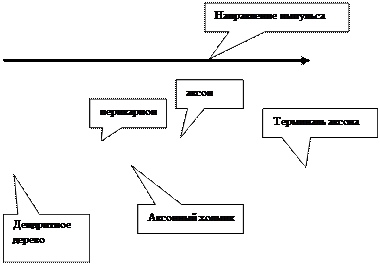 |
Структурно-функциональная схема нейрона.
Классификация нейронов. Существует несколько классификаций нервных клеток. В последние годы предпринимаются попытки создать единую комплексную классификацию.
Морфологическая классификация нейронов в соответствии с числом и типом отростков выделяет):
· мультиполярные - с множеством отростков,
· униполярные - с одним аксоном ,
· ложно- (псевдо-) униполярные – дендрит и аксон отходят от нейрона в виде одного отростка, а затем разделяются .
· биполярные - с одним аксоном и одним дендритом .
Морфологические классификации нейронов позволяют выделять и другие признаки клеток. Так, имеется классификация нервных клеток по размерам. Они делятся на гигантоклеточные, крупноклеточные, среднеклеточные, мелкоклеточные. По длине нейритов можно видеть длинноаксонные, среднеаксонные, короткоаксонные. По степени развития дендритного дерева мультиполярные нейроны бывают густоветвистые, средневетвистые и маловетвистые (изодендритные). Кроме этого можно подразделить нервные клетки по форме (звездчатые, округлые, грушевидные, пирамидные, веретеновидные, корзинчатые и т.д.), по медиатору характерному для них (адренэргические, холинэргические, глицинэргические, дофаминергические и т.д.).




Морфологические типы нейронов
Функциональная классификация нейронов позволяет выделить среди них:
· Афферентные нейроны (воспринимают информацию и передают ее в нервные центры). Их можно рассматривать как клетки, осуществляющие центростремительную передачу возбуждения. Данные клетки могут рассматриваться как чувствительные нейроны. Но, если понимать выражение чувствительность, в смысле того, что принимается и анализируется ЦНС, то не все клетки этой группы имеют в ней представительство. Так, афферентные нейроны интрамуральных (внутриорганных) нервных узлов ограничивают передачу возбуждения другими нервными клетками узла. Типичными чувствительными являются ложноуниполярные и биполярные нейроны спинномозговых и черепно-мозговых узлов.
· Ассоциативные нейроны, осуществляющие взаимодействие между нейронами. Являются основной популяцией нейронов в центральной нервной системе и по численности составляют абсолютное большинство. Типичным для них является мультиполярность, средние или мелкие размеры перикарионов.
· Эфферентные (центробежные) нейроны генерируют нервный импульс и передают возбуждение другим нейронам или клеткам других видов тканей. Значительная часть этих клеток контролируют мышцы и, таким образом, являются двигательными, обеспечивая произвольные и непроизвольные сокращения. Другие клетки формируют окончания на телах секреторных клеток и регулируют инкреторные, секреторные и экскреторные функции. Некоторые из нейронов направляют аксоны к кровеносным капиллярам выделяют инкрет в кровь, являясь нейросекреторными.
Среди функциональных типов нейронов выделяются возбуждающие, которые вызывают деполяризацию постсинаптических мембран клеток приводя к формированию так называемого потенциала действия, вызывающего стимулирующие эффекты. Тормозные нейроны приводят к гиперполяризации мембраны контролируемой клетки, снижая ее чувствительность к специфическим воздействиям. В этой связи, необходимо упомянуть о так называемых пейсмекерных нейронах. Эти клетки способны к автономному формированию возбуждения, независимо от внешних воздействий. Эти клетки характеризуются снижением мембранного потенциала покоя до порогового уровня даже без значительных внешних влияний или под действием возвратного возбуждения, возникающего в цепи нейронов. В последнем случае сигнал возвращается к возбуждающей клетке через цепь вставочных нейронов, что приводит к повторному формированию возбуждения по всей цепи. Часто клетка не может поддерживать постоянный потенциал покоя, и он опускается до минимального предела (порогового уровня) и происходит деполяризация мембраны с возбуждением нейрона.
Регенераторные процессы. Важной особенностью основной популяции зрелых нейронов является способность лишь к внутриклеточной регенерации. Они являются стабильной популяцией клеток и в обычных условиях не делятся. Но имеются исключения. Так, доказана способность к делению у нервных клеток в эпителии обонятельного анализатора, в ганглиях (скоплениях нейронов периферической нервной системы), контролирующих звуковую активность певчих птиц (g.vocalae). Некоторые авторы приписывают подобные возможности части интрамуральных (внутриорганных) вегетативных ганглиев (во всяком случае доказана их способность к неполному амитозу и митозу с формированием многоядерных полиплоидных нейронов). Предполагают возможность деления нейронов при наследственно передающемся заболевании старческого слабоумия (болезни Альцгеймера) и даже деление нейронов в пределах субвентрикулярных зон (вблизи желудочков мозга). Но вцелом, деление нервных клеток, как уже упоминалось, невозможно. Невозможность деления нейронов необходима для сохранения стабильности важнейших функциональных и макроанатомических межнейронных взаимодействий, что, в частности, позволяет нам сохранять индивидуальные особенности личности.
Невозможность клеточного деления, вовсе не предполагает неспособности нервных клеток к регенерации вообще. Нервные клетки на внутриклеточном уровне не только регенерируют, но характеризуются очень высокой физиологической внутриклеточной регенераторной способностью. Непрерывно происходит замещение старых органелл, белковых, липопротеидных и гликопротеидных комплексов (в частности, мембран), ресинтез ферментов, медиаторов и т.д. Это сопровождается высоким уровнем энергетического обмена, особенно в теле нейрона. В культуре нервной ткани нейроны активно делятся митозом.
Строение нейрона. На светооптическом уровне при общих методах окрашивания тела нервных клеток имеют оксифильную цитоплазму, крупное ядро округлой или овальной формы. Ядро занимает центральное положение, но иногда смещается к одному из полюсов нейрона, что чаще всего связано с реактивными процессами. В ядре хорошо развито одно или несколько ядрышек. В части нейронов можно видеть два и более ядер (до 10-15). Это более характерно для вегетативных узлов, особенно встроенных в структуру внутренних органов (внутриорганные или интрамуральные ганглии). Такие многоядерные клетки по сути являются редуцированными проявлениями деления не завершающихся полноценным цитокинезом. Следует иметь в виду, что у некоторых разновидностей нервных клеток можно видеть полиплоидный набор хромосом, что связано с эндомитозом. Полиплоидия облегчает высокую метаболическую активность нейронов. Кариоплазма отличается преобладанием диффузного (слабо конденсированного) хроматина. Нейроны имеют высокое сродство к солям серебра (аргирофильность). Специфичными для нейрона структурами цитоплазмы на светооптическом уровне являются хроматофильное вещество цитоплазмы и нейрофибриллы. Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при окрашивании нервных клеток основными красителями (метиленовым синим, толуидиновым синим, гематоксилином и т.д.) в виде зернистости. Зернистость может быть в виде крупных глыбок неправильной формы, а также в виде сети или мелкой зернистости. Это зависит от типа нейрона (крупные нейроны обычно имеют более крупные глыбки) и от его функционального состояния. На электроннооптическом уровне хроматофильное вещество цитоплазмы есть не что иное, как скопления цистерн зернистой эндоплазматической сети. Эти органеллы отсутствуют в аксоне и в аксонном холмике, но имеются в начальных сегментах дендритов. На светооптическом уровне также тигроид не виден в начале аксона, но прослеживается в дендритах, что позволяет идентифицировать вид отростков. Процесс разрушения или распада глыбок базофильного вещества называется тигролизом и наблюдается при реактивных изменениях нейронов (например, при их повреждении) или при их дегенерации. Тигролиз нередко сопровождается вакуолизацией цитоплазмы, Уплощенные цистерны ЭПС разбухают, а цитоплазма приобретает вакуолярную орагнизацию.
Нейрофибриллы, выявляют при импрегнации нейронов, в результате осаждения осадка серебра на структурах цитоскелета нейрона. Основой для выявления нейрофибрилл являются нейрофиламенты и нейротубулы, формирующие каркас нервной клетки.
Кроме этого, в нейронах довольно часто можно видеть липидные включения (зерна липофусцина). Они характерны для старческого возраста и часто появляются при дистрофических процессах. Зерна липофусцина являются остаточными тельцами, возникающими в результате неполного переваривания. Их накопление может приводить к нарушению нормальных метаболических процессов в клетках и их гибели.
У некоторых нейронов в норме обнаруживаются пигментные включения (например, с меланином), что обуславливает окрашивание нервных центров, содержащих подобные клетки (черная субстанция, голубоватое пятно).
Электроннооптическое строение и некоторые цитофизиологические особенности тела нейрона и его отростков. Несмотря на крайнее морфологическое разнообразие нейронов, они имеют ряд общих черт построения. Ядра нейронов, особенно крупноклеточных, имеют округлую форму. Кариолемма часто формирует впячивания, что может значительно увеличивать площадь контакта поверхности ядра с цитоплазмой (нейроплазмой). Ядерная оболочка имеет большое количество ядерных пор, что указывает на активные процессы обмена, в том числе РНК и субъединицами рибосом. Кариоплазма в крупных нейронах в основном светлая, но особенно в мелких, можно видеть и повышенную склонность к импрегнации (осмиофильность). Данные особенности на светооптическом уровне проявляются в гипохромности или гиперхромности ядер (т.е., пониженной или повышенной склонности к окрашиванию ядерными красителями). Хорошо развит ядрышковый аппарат. В ядре обычно имеется 1-2 крупных умеренной плотности ядрышка, занимающих центральное положение. В мелких нервных клетках ядрышки мельче, их может быть до 3-6 и более. При реактивных проявлениях в клетке можно наблюдать смещение ядрышка и его распад.
Матрикс цитоплазмы (нейроплазмы) гомогенный или мелкозернистый, слабой или умеренной электронной плотности. В нейроне сильно развита гранулярная эндоплазматическая сеть (ЭПС), представленная скоплениями или диффузно расположенными плоскими цистернами и трубочками. Как уже указывалось выше, ЭПС преобладает в теле, и может содержаться в начальных сегментах дендритов. За ней закреплено участие в процессах синтеза медиаторов и модуляторов, мембранных белков и т.д. Кроме связанных имеется и значительное число свободных полисом и рибосом.
Митохондрии в нейронах многочисленны. Они имеют средние и крупные размеры (диаметр 1-3 мкм), могут быть овальной или нитчатой формы. Кристы имеют трабекулярное строение. Нейроны в энергетическом отношении крайне зависимы от аэробного фосфорилирования и во взрослом состоянии фактически не способы к анаэробному гликолизу. В связи с этим нервные клетки находятся в выраженной зависимости от поступления кислорода и глюкозы и при нарушении кровотока нервные клетки практически сразу прекращают свою жизнедеятельность. Момент прекращения кровотока в головном мозге означает начало клинической смерти. Практически сразу же начинаются процессы саморазрушения в нейронах, и прекращается их специфическая функциональная активность. Их мембраны деполяризуются. Митохондрии, ЭПС, ядерные оболочки набухают, а затем и разрушаются. Начинаются процессы аутолиза и перекисного окисления. Эти процессы могут носить обратимый или необратимый характер. При мгновенной смерти, при комнатной температуре, и нормальной температуре тела процессы саморазрушения в нейронах обратимы в течение 5-7 минут. Это и является сроком так называемой клинической смерти, когда возможно оживление организма. Необратимые изменения в нервной ткани приводят к переходу от клинической смерти к биологической. В случае продолжительного умирания, высокой внутренней или внешней температуре, особенно при сердечно-сосудистой и дыхательной недостаточности время необратимого разрушения нервных клеток значительно сокращается до 1-2 и менее минут. Наоборот, внезапная остановка сердечной деятельности при низких температурах окружающей среды и головного мозга может значительно удлинять время клинической смерти.
В нейронах значительного развития достигает пластинчатый комплекс. Он может располагаться компактно или быть рассеян в цитоплазме тела нейрона. Нередко наблюдается поляризация этой органеллы. Это указывает на преимущественное направление транспортных потоков и процессов созревания секретируемых продуктов (медиаторов) от тела к аксону.
Специфическими органеллами нейрона считают нейрофиламенты и нейротубулы. Нейрофиламенты представляют собой промежуточные филаменты диаметром 8-10 нм, образованные фибриллярными белками. Основной функцией этих элементов цитоскелета является опорная - для обеспечение стабильной формы нейрона. Подобную же роль играют тонкие микрофиламенты (поперечный диаметр 6-8 нм), содержащие белки актины. В отличие от подобных микрофиламентов в других тканях и клетках, они не соединяются с микромиозинами, что делает невозможным активные сократительные функции в зрелых нервных клетках.
Нейротубулы по основным принципам своего строения фактически не отличаются от микротрубочек. Они, как и все микротрубочки имеют поперечный диаметр около 24 нм, кольца замыкают 13 молекул глобулярного белка тубулина. Микротрубки полярны. В нервной ткани они выполняют очень важную, если не сказать уникальную роль. Как и всюду они несут каркасную (опорную) функцию. В отличие от нейрофиламентов являются менее стабильными и обеспечивают процессы циклоза. Именно полярность микротрубки, в которой имеется отрицательно и положительно заряженные концы, позволяет контролировать диффузионно-транспортные потоки в аксоне (так называемый быстрый и медленный аксоток). Их подробное описание приведем несколько ниже.
В цитоплазме тел нейронов часто встречаются лизосомы. Они участвуют в пластических процессах, осуществляя катаболизм (разрушение) старых органелл и структур. В результате переваривания образуются остаточные тельца. Часть из них содержит непереваренные остатки органелл, включающие липофусцин. Избыточное накопление липофусцина может приводить к дистрофическим процессам в нейроне, к нарушению его специфической активности и даже гибели. Такие явления характерны для старческих изменений и при различных патологических воздействиях.
В теле нейронов можно видеть также транспортные пузырьки, часть из которых содержит медиаторы и модуляторы. Они окружены мембраной. Их размеры и строение зависят от содержания того или иного вещества.
Особенности морфологии отростков нейронов. Дендриты короткие отростки, нередко сильно ветвятся. Их ветвления более выражены в терминальных областях. Дендриты в начальных сегментах содержат органеллы подобно телу нейрона. Хорошо развит цитоскелет.
Аксон чаще всего длинный, слабо ветвится или не ветвится. Ветвления чаще всего наблюдаются в терминальных зонах. Уже в начальном сегменте аксона, в нем отсутствует гранулярная ЭПС. Микротрубочки и микрофиламенты располагаются упорядочено. В цитоплазме аксона видны митохондрии, транспортные пузырьки. Аксоны в основном миелинизированы и окружены отростками олигодендроцитов в центральной нервной системе, или леммоцитами в периферической нервной системе. Начальный сегмент аксона нередко расширен и имеет название аксонного холмика. Именно в зоне аксонного холмика происходит суммация поступающих в нервную клетку сигналов и если возбуждающие сигналы достаточной интенсивности, то в аксоне формируется потенциал действия и возбуждение направляется вдоль аксона, передаваясь на другие клетки (потенциал действия).
От отростков нейронов, а нередко и от его тела, отходят небольшие выпячивания, которые имеют форму напоминающую шипики, откуда и получили название. Особенно развиты шипики на некоторых нервных клетках центральной нервной системы. Шипики являются постсинаптическими структурами и соответствуют зонам взаимодействия одних нервных клеток с другими. Они имеют элементы цитоскелета, митохондрии. Нередко видны уплощенные цистерны и электронно-плотное вещество мембраны.
Аксоток (аксоплазматический транспорт веществ). Нервные волокна имеют своеобразный структурный аппарат– микротрубочки, по которым перемещаются вещества от тела клетки на периферию (антероградный аксоток) и от периферии к центру (ретроградный аксоток). Направление аксотока обеспечивает полярность микротрубочек. В нем участвует белок кинезин, взаимодействующий с тубулином микротрубочек, и осуществляющий транспорт с затратой энергии АТФ.
Различают быстрый (со скоростью 100-1000 мм/сут.) и медленный (со скоростью 1-10 мм/сут.) аксоток.
Быстрый аксоток - одинаков для различных волокон; требует значительной концентрации АТФ; происходит с участием транспортных пузырьков. Он осуществляет транспорт медиаторов и модуляторов.
Медленный аксоток - за счет него от центра к периферии распространяются биологически активные вещества, а также составляющие компоненты мембран клеток и белков.
За счет антероградного тока происходит дифференциация мышц. Это имеет большое биологическое значение. Есть мышцы фазные (мышцы конечностей) и тонические (поддерживают позу). В эксперименте установлено, что если перерезать нервы, иннервирующие эти мышцы, а затем произвести перекрест иннервации, то есть центральный конец нерва, иннервирующего фазные мышцы, подшить к нерву, иннервирующему тонические мышцы, то после прорастания нервов, фазные мышцы начинают выполнять функции тонических, а тонические - фазных. Меняется их структура, так как за счет аксотока обеспечивается трофическая функция двигательных нервов. За счет ретроградного тока нейротропные вещества поступают от периферии к центру, оказывая трофическое влияние на саму нервную клетку. За счет ретроградного тока в ЦНС могут поступать различные токсические вещества.
Синапсы. Синапсы это специфические контакты нейронов обеспечивающие передачу возбуждения от одной нервной клетки к другой. В зависимости от способов передачи возбуждения (так называемого потенциала действия) выделяют химические (рис.25). и электрические синапсы.
Эволюционно более древними и примитивными являются электрические синаптические контакты. Они по строению близки к щелевидным контактам (нексусам) и представляют собой ограниченные области контактов двух соседних нейронов, где расстояние между соседними мембранами сужено до 3-4 нм. В зоне этих сужений имеются белковые мембранные комплексы аналогичные коннексонам. Коннексоны представляют собой группу высокомолекулярных интегральных белков. В центральной зоне они формируют пору. При этом белки соседних мембран тесно взаимодействуют между собой, что приводит к тому, что пора, формируемая одной клеткой, переходит на соседнюю клетку. Через нее могут свободно диффундировать вода, ионы, низкомолекулярные органические вещества (в том числе и гормоноподобные). Считается, что обмен происходит в обе стороны, но имеются случаи, когда возбуждение передаются в одном направлении.
Таким образом, возбуждение в таком синапсе в большинстве случаев идет в обе стороны и носит равновероятностный, диффузный характер. Такие контакты часто встречаются у низших беспозвоночных и хордовых. У млекопитающих электрические контакты имеют большое значение в процессе межнейронных взаимодействий в эмбриональном периоде развития. С дифференцировкой нервной ткани их число убывает. Подобный вид контактов у взрослых млекопитающих имеет место в ограниченных участках, например их можно видеть в мезэнцефалическом ядре тройничного нерва.
Химические синапсы. Химические синапсы для передачи возбуждения от одной нервной клетки к другой используют специальные вещества – медиаторы, от чего и получили свое название. Кроме медиаторов ими используются и модуляторы. Модуляторы это специальные химические вещества, которые сами возбуждения не вызывают, но могут либо усиливать, либо ослаблять чувствительность к медиаторам (то есть модулировать пороговую чувствительность клетки к возбуждению). Согласно закону Дейля каждый нейрон содержит один специфичный для него медиатор и один или несколько модуляторов. Эти вещества накапливаются в синаптических пузырьках, которые отделены от матрикса цитоплазмы одной двухслойной мембраной. Форма, размеры, строение пузырьков зависит от содержащегося в них вещества.
Любой из химических синапсов обеспечивает однонаправленную передачу возбуждения. В связи с чем в химическом синапсе выделяют следующие зоны:
· Пресинаптическая зона (пресинаптическое расширение, наиболее часто представляющее собой терминаль аксона)
· Синаптическая щель (размерыетром 20-50 нм)
· Постсинаптическая зона
Пресинаптическая зона всегда содержит синаптические пузырьки, элементы цитоскелета (нейротубулы и нейрофиламенты), митохондрии. Часто мембрана, обеспечивающая передачу импульса имеет хорошо выраженное электронноплотное вещество. Оно представляет собой скопление мембранных белков, одной из функций которых является направленное выделение содержимого синаптических пузырьков в синаптическую щель. Таким образом, выделение медиаторов осуществляется лишь через строго определенные участки пресинаптической мембраны.
Синаптическая щель изолируется от окружающего межклеточного вещества нейроглией (в центральной нервной системе отростками астроцитов), принимает медиаторы из пресинаптической зоны
Постсинаптическая зона включает электронноплотное вещество постсинаптической мембраны, митохондрии. Электронноплотное вещество представлено комплексом мембранных белков, в том числе рецепторных (на гликокаликсе), белков ионных каналов, и ферментами расщепляющими медиаторы.
В зависимости от того, какие структуры взаимодействуют в синапсе, можно выделить:
- аксо-дендритические (пресинаптическая структура аксон, постсинаптическая - дендрит),
- аксо-аксональные (аксон с аксоном)
- аксо-соматические (аксон с телом нервной клетки).
В нервной системе имеются и иные синаптические взаимодействия, в том числе дендро-дендритические, дендро-соматические и т.д.
По строению выделяют сложные и простые синапсы. Простые синапсы содержат лишь одну поверхность взаимодействия между терминалью и посттерминальным (постсинаптическим) образованием. Сложные синапсы имеют несколько поверхностей взаимодействия. Они могут быть дивергентного, конвергентного, параллельного или эфаптического типа. Дивергентный синапс, это окончание, когда одно пресинаптическое расширение обеспечивает передачу возбуждения к нескольким постсинаптическим структурам. Конвергентный синапс предполагает одну постсинаптическую зону, к которой импульс передается от нескольких пресинаптических расширений. Параллельный синапс предполагает возможность передачи возбуждения от одной нервной структуры к близлежащей после ее стимуляции соседним претерминальным образованием. В центральной нервной системе преобладающими являются сложные синапсы.
Функционально выделяют возбуждающие и тормозные синапсы. Возбуждающие приводят к деполяризации постсинаптической мембраны, что приводит к активации нервной клетки и возникновению в ней потенциала действия. Тормозные, наоборот, приводят к гиперполяризации мембраны, что снижает пороговую чувствительность нейрона к внешним влияниям.
Синапсы могут быть симметричными (когда электронноплотное вещество хорошо выражено как на синаптической, так и постсинаптической мембранах) и асимметричными (слабее выявляется электронноплотное вещество пресинаптической мембраны). Симметричным синапсам часть авторов приписывает роль тормозных, а асимметричным – возбуждающих.
По основному медиатору, содержащемуся в синаптических пузырьках, синапсы делятся на группы:
1. Холинергические (ацетилхолинергические). Возбуждающие и тормозные.
2. Адренергические (моноаминергические, норадренергические, дофаминергические). В основном, возбуждающие, но есть и тормозные.
3. Серотонинергические (иногда приписываются к предыдущей группе). Возбуждающие.
4. ГАМК-ергические (медиатор гаммааминомаслянная кислота). Тормозные.
5. Глутаматергические. Возбуждающие.
6. Аспартатергические. Возбуждающие.
7. Глицинергические (тормозные).
8. Пептидергические. Большая группа вешеств, в основном в ЦНС. Это может быть вазоинтерстициальный полипептид, вазопрессин, вещество Р (медиатор боли), нейропептид Y, окситоцин, бета-эндорфин и энкефалины (противоболевые), динорфин и т.д.
В последние годы значительный интерес проявляется к роли закиси азота как к медиатору, в том числе и внесинаптической нервной передачи. В частности, значительная группа нейронов-водителей ритма (пейсмекеров) имеет NO-активность.
В зависимости от типа медиатора синаптические пузырьки имеют разнообразную структуру. Все они отделены от гиалоплазмы одной двухслойной мембраной. Холинсодержащие пузырьки электронносветлые, диаметром 40-60 мкм. Адренсодержащие - с электронноплотной сердцевиной, светлой каемкой, диаметром 50-80 мкм. Глицинсодержащие и ГАМК-содержащие – имеют овальную форму. Пептидсодержащие – с электронноплотной сердцевиной, светлой каемкой, диаметром 90-120 мкм.
Механизм передачи возбуждения в химическом синапсе: Импульс, приходящий по афферентному волокну, вызывает возбуждение в синаптической бляшке и приводит к выделению медиатора через пресинаптическую мембрану. В периферических синапсах медиатором служит преимущественно ацетилхолин. В синаптическую щель поступает медиатор. На пресинаптической мембране имеются активные зоны, то есть участки пресинаптической мембраны, где больше всего происходит выброс ацетилхолина в синаптическую щель. Рядом расположены неактивные участки. На постсинаптической мембране имеются специальные холинорецепторы (для медиатора ацетилхолина) или адренорецепторы (для норадреналина).
Виды секреции ацетилхолина:
1) квантовая секреция (вызванная) - с помощью импульса - является основным фактором, вызывающим деполяризацию постсинаптической мембраны и сокращение мышц.
2) неквантовая секреция - на нее приходится около 90% выделяемого медиатора, но, т.к. он поступает микродозами, не сопровождается деполяризацией постсинаптической мембраны, и не вызывает сокращения мышц. Этот механизм участвует в ауторегуляции секреции ацетилхолина и обеспечивает трофические воздействия на мышцу
После выхода через активные зоны медиатор взаимодействует с холинорецепторами постсинаптической мембраны. Происходит изменение концентраций Na+ и К+, что приводит к возникновению потенциала действия. В постсинаптическом волокне медиатор холинэстераза разрушает ацетилхолин на ацетатную группу (утилизируется) и холин, который возвращается в постсинаптическую бляшку и участвует в ресинтезе медиатора.
Особенности функционирования химического синапса:
1. Односторонняя передача возбуждения (от нерва на рабочий орган) – связано с тем, что рецепторы, воспринимающие медиатор, находятся на постсинаптической мембране.
2. “Синаптическая задержка” – время, необходимое на выделение медиатора в пресинаптической бляшке.
3. Следовые явления или синаптическое последействие – после прекращения раздражения продолжают поступать импульсы, т.е. продолжается потенциал действия (связано с тем, что необходимо время для разрушения медиатора холинэстеразой).
4. Суммация возбуждения – при действии подпорогового раздражителя выделяется небольшое количество медиатора, способное вызвать миниатюрный потенциал (т.е. локальное возбуждение), который не передается на постсинаптическую мембрану. Суммация может быть последовательная (временная) и пространственная. Последовательная - при частом поступлении по одному пути импульсов подпороговой величины происходит накопление достаточного количества медиатора и возникает возбуждение (потенциал действия), которое передается на постсинаптическую мембрану. Пространственная - при раздражении 2-х рецептивных полей, расположенных рядом повышается возбудимость и может возникнуть ответная реакиця.
5. Трансформация ритма раздражения в сторону увеличения или уменьшения количества импульсов на выходе – связано с низкой функциональной лабильностью синапса.
6. Высокая чувствительность к химическим веществам и специфичность (рецепторы обладают избирательной чувствительностью к медиаторам.
7. Быстрое утомление – связано с низкой функциональной лабильностью синапса и ограниченным наличием медиатора.
8. Способны изменять свою возбудимость в сторону повышения (синаптическая потенциация) или в сторону уменьшения (при длительном действии - десенситизация)
Цитофизиологические и нейрофизиологические основы межнейронной передачи. Возбуждение в нейроне, как уже указывалось, связано со специфическими или (и) неспецифическими воздействиями на нервную клетку, приводящими к деполяризации мембраны нейрона. Потенциал покоя нервной клетки составляет 60-70 мВ, скорость деполяризации 1-2 мс, время реполяризации 1-2 мс. В момент деполяризации возбуждение достигает аксонального холмика, где оно может затормаживаться (если не достигает достаточной силы – пороговой чувствительности) или может распространяться далее по аксону к пресинаптической структуре (терминали аксона). Когда деполяризационная волна возбуждения достигает пресинаптического расширения, в нем открываются кальциевые каналы, в результате чего концентрация ионов кальция увеличивается. Это способствует связыванию синаптических пузырьков с пресинаптической мембраной. В данном процессе участвуют микротрубочки. Белок синаптических пузырьков – синаптобревин связывается с белками пресинаптических мембран (синтаксин и SNAP-25), после чего мембраны сливаются и содержимое пузырьков с медиатором выделяется в синаптическую щель. Медиатор взаимодейтсвует с рецепторами постсинаптических мембран. Часть из них, взаимодействуя с ионными каналами, открывает К/Na каналы и приводит к деполяризации мембран и возбуждению следующей нервной клетки. Другие – активируют процессы выделения натрия из клетки и гиперполяризации мембран нервных клеток.
В последующем связь медиаторов с рецепторами разрывается. Медиатор либо метаболизируется, либо подвергается обратному всасыванию пресинаптическими мембранами, либо захватывается мембранами астроцитов с последующей передачей медиатора к нервным клеткам.
Нейроглия. Впервые данный термин ввел Вирхов для описания клеток между нейронами. Занимают они около половины всего объема ткани. К ней относят нейроглию центральной нервной системы и периферической нервной системы. Выделяют макроглию и микроглию
Среди макроглии центральной нервной системе имеются волокнистые (фиброзные) и протоплазматические астроциты, олигодендроциты, эпендимоциты (в том числе и танициты). В периферической нервной системе нейроглия представлена леммоцитами (шванновскими клетками) и сателлитоцитами.
Астроциты. Astro – звездчатые, cites – клетки. В дословном переводе звездчатые клетки. Названы так из-за обилия отростков, отходящих от тела клетки. Многочисленные отростки ветвятся и окружают другие структуры мозга. Астроциты есть только в ЦНС и анализаторах – производных нервной трубки. Среди них встречаются волокнистые и протоплазматические астроциты. Терминали отростков обеих типов клеток имеют пуговичные расширения (ножки астроцитов), значительная часть из которых заканчивается в периваскулярном пространстве, окружая капилляры периваскулярными глиальными бухтами занимающими 80% обменной поверхности микрососудов.
Волокнистые астроциты имеют многочисленные, длинные, тонкие, слабо или совсем не ветвящиеся отростки. В основном присутствуют в белом веществе мозга.
Протоплазматические астроциты отличаются короткими, толстыми и сильно ветвящимися отростками. Имеются преимущественно в сером веществе мозга. Морфология астроцитов отличается крайним разнообразием и коррелирует с формой капиллярных петель и нейронных ансамблей в ЦНС. Клетки диффузно распределены в объеме мозга. Их отростки взаимопереплетаются между собой, так и другими элементами нейропиля. Астроциты занимают исключительное положение в ЦНС, располагаясь между телами нейронов, немиелинизированной и миелинизированной частями нервных отростков, синапсами, кровеносными сосудами, подэпендимными пространствами изолируя и в то же время структурно связывая их.
Специфическим маркером астроцитов является так называемый глиальный фибриллярный кислый белок, из которого образуются промежуточные филаменты.
Строение астроцитов. Клетки имеют относительно крупные светлые ядра, со слабо развитым ядрышковым аппаратом. Цитоплазма слабо оксифильная. В ней слабо развиты гладкая и гранулярная ЭПС, пластинчатый комплекс. Митохондрий мало, они небольших размеров. Цитоскелет развит умеренно в протоплазматических и хорошо - в волокнистых астроцитах. Между клетками значительное число щелевидных и десмосомоподобных контактов.
В постнатальный период жизни человека астроциты способны к миграции, особенно в зоны повреждения и, как полагают, способны к пролиферации.
Функции астроцитов:
1.Участвуют в гематоэнцефалическом и ликворогематическом барьерах. Астроциты своими ножками покрывают капилляры, поверхности мозга и участвуют в транспорте веществ от сосудов к нейронам и наоборот. Способы передавать метаболиты нейронам.
2. Обеспечивают ионный обмен, особенно ионов калия. При активации нейронов, их частая и длительная деполяризация может вести к значительному увеличению ионов калия, что может изменять мембранный потенциал нейронов, повышая их чувствительность к внешним воздействиям. Астроциты, захватывая избыточный калий, предотвращают перевозбуждение. Нарушения в этой функции могут вести к развитию состояний эпилепсии.
3. Изолируют рецепторные поверхности тел нейронов и синапсов. При этом клетки способны к ритмичным сокращениям (по некоторым авторам к набуханию) изменяя зоны изоляции.
4.Способны к захвату нейромедиаторов (глиоксиловой кислоты, гамма-аминомаслянной кислоты) из зон синаптической передачи. Под их влиянием изменяется активность самих астроцитов, что приводит к внесинаптической модуляции сигнала в соседних нервных клетках и их отростках.
5.Фагоцитоз погибших нейронов. На месте фагоцитированных нейронов в результате их гибели, глиальные рубцы. Это скопления гипертрофированных астроцитов, заменяющих собой рубцы в периферических органах.
6.Выделяют большое количество биологически активных веществ (факторы роста нервов, факторы роста фибробластов, ангиогенные факторы, эпидермальный фактор роста, интерлейкин–I, простагландины), контролирующих местные межклеточные метаболические и внесинаптические информационные взаимодействия. Факторы роста способны инициировать и ускорять рост отростков нейронов. Выделение интерлейкина–I, способность экспрессировать МНС-комплексы 1 и 2 классов, указывает на роль астроцитов в формировании специфических иммунных реакций и антигенпрезентирующую функцию.
Выделение факторов роста фибробластов, компонентов межклеточного вещества (ламелин, фибронектин), простагландинов, антиваскулярных факторов позволяет астроцитам контролировать состояние местного кровотока в ЦНС.
7.В эмбриональном развитии человека предшественники астроцитов контролируют направление миграции астробластов, во всяком случае, в части зон головного мозга (мозжечок, гипоталамус), а также в зоне их отростков (зрительный нерв).
В постнатальном развитии стабилизируют структуры ЦНС, ингибируя рост отростков нейронов и в то же время, предотвращая апоптозы (запрограммированную гибель) нейронов, предотвращая избыточное снижение их числа при повреждении.
Олигодендроциты. Олигодендроциты (олигодендроглиоциты) центральной нервной системы относятся к миелинообразующим клеткам и участвуют в формировании нервных волокон. Подобную им функцию в периферической нервной системе играют леммоциты (шванновские клетки) периферической нервной системы.
Тела по размерам как правило мельче астроцитов. Ядра мелкие, округлые, темноокрашенные. Ядрышки мелкие. Отростков мало. Они тонкие, не ветвятся или слабо ветвятся и заканчиваются вокруг аксонов и дендритов нервных клеток. Часть олигодендроцитов концентрируется в непосредственной близости к телам нервных клеток (сателлитные олигодендроциты). Терминальная зона каждого отростка участвует в формировании сегмента нервного волокна, то есть каждый олигодендроцит обеспечивает окружение сразу нескольких нервных волокон. Цитоплазма слабоксифильная и при общих методах окрашивания сливается с нейропилем. На электроннооптическом уровне–в цитополазме хорошо развиты органеллы по составу близкие к нейронам, но в отличие от них не имеют развитого цитоскелета.
Леммоциты (шванновские клетки) периферической нервной системы. Окружают отростки нейронов в периферической нервной системе. При этом клетки имеют удлиненную форму, распластываясь на поверхности аксона, не имеют отростков и обеспечивают формирование глиальной оболочки в одном сегменте миелинового или безмиелинового нервного волокна.
В области формирования корешков спинномозговых и черепно-мозговых нервов формируют скопления (глиальные пробки), предотвращая проникновение отростков ассоциативных нейронов ЦНС за ее пределы.
Кроме леммоцитов в периферической нервной системе имеются сателлитные (мантийные) глиоциты в периферических нервных узлах вокруг тел нейронов, глиоциты нервных окончаний, конкретные морфологические особенности которых рассматриваются при изучении нервных окончаний и анатомии нервных узлов.
Леммоциты характеризуются удлиненными, темноокрашенными ядрами, слабо развитыми митохондриями и синтетическим аппаратом (гранулярная, гладкая ЭПС, пластинчатый комплекс).
Мантийные клетки уплощенной формы с несколькими отростками. Они распластаны на поверхности нейрона и формируют вокруг него глиальную капсулу. Ядра уплощены, темные, со слабо развитым ядрышковым аппаратом. Синтетический аппарат слабо развит. Клетки обеспечивают изолирующую, трофическую, опорную, защитную функции. Играют барьерную роль.
Функции олигодендроглии и леммоцитов.
1. Формируют оболочки вокруг нервных клеток, обеспечивая барьерно-транспортные функции и изоляцию тел нейронов.
2. Образуют миелиновые оболочки, участвуя в пров
| <== предыдущая страница | | | следующая страница ==> |
| ТЕМА: КОСТНЫЕ ТКАНИ | | |
Дата добавления: 2014-09-10; просмотров: 828; Нарушение авторских прав

Мы поможем в написании ваших работ!