Главная страница Случайная лекция

Мы поможем в написании ваших работ!
Порталы:
БиологияВойнаГеографияИнформатикаИскусствоИсторияКультураЛингвистикаМатематикаМедицинаОхрана трудаПолитикаПравоПсихологияРелигияТехникаФизикаФилософияЭкономика

Мы поможем в написании ваших работ!
Данные многолетней динамики численности популяций лесных насекомых и их представление
Сезонная динамика численности популяции. Таблицы выживаемости
1 2
Рисунок 6.1 - Схема расположения анального и полового отверстий у куколок бабочек (1- самец, 2- самка; а – анальное отверстие, п – половое отверстие)
Половой индекс популяции насекомых может варьировать в широких пределах. У насекомых, дающих вспышки массового размножения, на начальных этапах градации самки могут значительно доминировать, а при затухании вспышки – их количество в популяции становится минимальным. Соотношение полов изменяется в значительном диапазоне в популяциях насекомых с повышенной миграционной активностью, например, у сибирского шелкопряда.
Масса особей является одним из наиболее важных качественных показателей, характеризующим состояние популяции насекомого. Как правило, у бабочек самки по массе больше самцов. Масса особей обуславливается количеством и качеством корма, используемым личинкой. При анализе весовых параметров особей можно определять массу яйцекладок, личинок разных возрастов, куколок и имаго. Однако, как правило, определяют массу куколок. Это связано с тем, что по массе куколки можно оценить потенциальную плодовитость насекомых в данной популяции. Такая оценка возможна потому, что масса куколок-самок хорошо коррелирует с массой имаго, а масса имаго, в свою очередь, с плодовитостью бабочек, вышедших из куколок.
Придержки плодовитости различных видов насекомых в зависимости от массы их куколок или коконов приводятся в книге «Надзор, учет и прогноз массовых размножений хвое- и листогрызущих насекомых в лесах СССР» (1965). Однако, как показывает опыт, для сибирских популяций многих видов насекомых необходимо делать некоторую поправку в сторону уменьшения плодовитости.
Масса особей определяется поштучным взвешиванием куколок или коконов самок и самцов отдельно. Далее рассчитываются статистические показатели массы особей в популяции – средняя масса m самцов и самок, стандартная ошибка средней массы sm – по формулам:
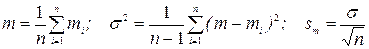 , (6.8)
, (6.8)
где mi - масса особей (самцов или самок) в выборке; σ2 и σ - соответственно дисперсия и стандартное отклонение от среднего; sm - стандартная ошибка среднего; n - число особей в выборке.
Кроме того, определяется размах значений массы самцов и самок – минимальные и максимальные значения масс самок и самцов.
Для более объективной оценки популяции насекомого по показателям массы особей необходимо проанализировать распределение особей по классам массы и долю особей с различной массой в популяции. Распределение куколок по массе будет значительно отличаться как на различных фазах градации, так и в различных частях очага массового размножения.
В связи с тем, что масса особей зависит от качества и объема доступного корма в период основного питания насекомого, с увеличением плотности популяции насекомого наблюдается снижение массы особей
Если анализировать изменение массы особей в процессе развития вспышки массового размножения насекомого, то, как правило, наблюдаются следующие закономерности. В межвспышечный период плотность популяции насекомых крайне мала, однако, масса особей достаточно велика. С ростом численности популяции, начиная со второй половины фазы максимума численности, происходит выраженное уменьшение массы особей. Резко снижается масса особей насекомых вспышечного вида после проведения лесозащитных мероприятий.
Плодовитость самок непосредственно связана с их массой и также является одним из важнейших качественных популяционных показателей, изменяющихся в широких пределах.
Оценка плодовитости насекомых может проводиться несколькими способами:
1. Подсчет яиц в кладке или взвешивание собранных кладок. Подсчет яиц или взвешивание собранных яйцекладок можно проводить для видов, откладывающих яйца полностью в одну кладку (например, у непарного шелкопряда). В случае взвешивания кладок, далее производится установление количества яиц в кладке по существующим таблицам (Надзор…, 1965). Однако лучше первоначально выявить взаимосвязь массы кладки и количества яиц, содержащихся в ней, для конкретного района исследования, а в дальнейшем уже пользоваться выявленной связью для определения плодовитости бабочек по массе яйцекладок.
2. Определение плодовитости бабочек методом их препарирования и непосредственного подсчета количества яиц в яйцевых трубках.
3. Взвешивание куколок – самок (или коконов) насекомых и дальнейшее определение плодовитости по имеющимся таблицам (Надзор…., 1965). Однако, как уже отмечалось, желательно первоначально выявить связь, существующую между массой куколок и плодовитостью имаго в конкретном районе исследований, а в дальнейшем уже использовать ее для определения плодовитости особей.
Следует отметить, что в различных частях ареала у одного вида насекомого связь между плодовитостью имаго и массой куколок может значительно изменяться.
В процессе развития вспышки массового размножения с изменением массы особей изменяется и плодовитость самок. На начальных этапах подъема численности вида масса особей и их плодовитость максимальны. На фазе депрессии масса особей и их плодовитость достигают своих минимальных значений (таблица 6. 1).
Таблица 6.1 - Масса куколок-самок и плодовитость монашенки на разных фазах вспышки массового размножения (цит. по « Надзор, учет и прогноз…..», 1965)
| Фаза градации* | Масса куколок, г | Плодовитость бабочек, яиц | ||||
| max | средняя | min | max | средняя | min | |
| Фаза роста численности | 0,9 – 1,0 | 0,7-0,8 | – | 600-630 | 420-520 | – |
| Собственно вспышка | – | 0,5-0,6 | – | – | 250-350 | – |
| Фаза кризиса | – | 0,3-0,4 | 0,2 | – | 100-170 | 0-20 |
* Фазы градации приводятся по « Надзор, учет и прогноз…..», 1965.
Следует подчеркнуть, что если в научных целях для оценки состояния популяции насекомого определяют как массу особей, так и плодовитость бабочек, то в практических целях часто определяют лишь массу особей.
Смертность от энтомофагов и болезней.Уровеньзараженности насекомых паразитами, болезнями и гибель их от хищных насекомых в процессе развития вспышки массового размножения закономерно изменяется. В связи с этим смертность от энтомофагов и болезней является объективным показателем состояния популяции, который необходимо учитывать при назначении лесозащитных мероприятий.
Болезни насекомыхмогут вызываться различными микроорганизмами – бактериями (бактериозы), вирусами (вирозы), грибами (микозы). Некоторые болезни насекомых обусловливаются действием одноклеточных животных (протозоонозы). Особую группу составляют болезни, вызываемые нематодами и трематодами (гельминтозы).
Зная болезни, свойственные определенным насекомым, и условия, при которых болезни переходят в эпизоотии, можно заблаговременно прогнозировать снижение численности насекомого-вредителя. В связи с этим выявление болезней насекомых является необходимым этапом при разработке прогнозов их массового размножения.
Для многих вредителей леса (например, для сибирского шелкопряда, непарного шелкопряда, монашенки, пилильщиков, пядениц) характерны вирусные заболевания. По современной классификации вирусы, поражающие насекомых, представлены в 13 семействах (Патогены насекомых…, 2001). Наиболее изученными являются вирусы, относящиеся к сем. Baculoviridae (бакуловирусы), которые включают в себя группу ядерных полиэдрозов и гранулезов. Эти заболевания известны только для насекомых и названы так благодаря форме белковых включений (полиэдров и гранул), образующихся в инфицированных клетках.
Характерной биологической особенностью бакуловирусов является их способность к латенции. Эти вирусы могут в течение ряда генераций насекомого находиться в скрытой форме, передаваясь трансовариально (через яички), а затем переходить в острую форму болезни, вызывая, как правило, гибель в короткие сроки большей части популяции насекомых. Вирозы носят резко выраженный эпизоотический характер и приводят к вымиранию популяций с последующим длительным отсутствием вредителя. Вирозам подвержены такие вредители леса как непарный шелкопряд, монашенка, боярышница, сосновая пяденица, ивовая волнянка, сосновая совка и многие другие.
Бактериальные болезни могут быть причиной значительной смертности насекомых в природе, хотя они редко носят такой массовый характер, как вирусные и некоторые грибные болезни. Патогенные для насекомых бактерии относятся к порядкам Bacillales, Clostridiales, Eubacteriales и Pseudomonodales (Патогены насекомых…, 2001). Практический же интерес представляют некоторые споровые и бесспоровые бактерии. Ведущая роль среди возбудителей бактериальных болезней насекомых принадлежит Bacillus thuringiensis. Особенностью этих бактерий является образование одновременно со спорой кристаллического включения, которое содержит токсическое вещество. Патогенное действие этих бактерий проявляется в параличе, наступающем вскоре после попадания бактериальных спор и клеточных включений в кишечник насекомого. Парализованные насекомые перестают питаться и гибнут.
В природе часто наблюдается гибель насекомых от красного бактериоза, вызываемого бесспоровой бактерией Serratia marcescens. Из бесспоровых бактерий, патогенных для многих насекомых, следует отметить также представителей рода Pseudomonas.
Среди возбудителей микозов насекомых одной из важнейших групп являются энтомофторовые грибы (Zygomycetes, Entomophtorales). Большинство представителей этих грибов, являясь облигатными паразитами членистоногих, способны в короткий срок и на обширных территориях вызывать массовые заболевания и выступать в качестве одного из главнейших механизмов регуляции численности своих хозяев.
Болезни этого типа называются энтомофторозами. Пораженные энтомофторозом насекомые мумифицируются, их ткани заполняются плотным сплетением толстого одноклеточного мицелия. Энтомофторозы часто поражают сосновую совку, кольчатого шелкопряда и других насекомых.
Другой группой грибных болезней являются мускардинозы, которые вызываются энтомопатогенными гифомицетами. Мускардинозы бывают различного типа: белый, зеленый, розовый, красный. При мускардинозе тело насекомых мумифицируется, поверхность его часто покрывается налетом гриба, имеющим различную окраску. Течение этих болезней зависит от экологических условий и физиологического состояния вредителя. Эти микозы часто поражают сосновую пяденицу, сосновую совку, многих пилильщиков, звездчатого ткача и др.
В ограничении численности зимней пяденицы, непарного шелкопряда, златогузки, краснохвоста, некоторых листоверток значительную роль играют болезни, вызываемые простейшими. В отличие от большинства возбудителей, простейшие передаются от зараженных насекомых в следующее поколение. Заражение происходит с пищей. Эти болезни носят хронический характер, ослабляя популяцию насекомого и вызывая различные аномалии.
Заболевания насекомых, вызываемые энтомогельминтами, изучены в меньшей степени. Среди энтомогельминтов наибольшее значение имеют нематоды, паразитирующие в полости тела или в тканях насекомых. В теле насекомого может быть обнаружено до 1500 штук нематод. Нематоды могут поражать все фазы развития насекомого – от яйца до имаго. Насекомые, пораженные гельминтами, съеживаются и разрушаются, у живых пораженных насекомых отмечается беспокойство и судорожные движения. Нематоды могут выступать и как переносчики патогенных бактерий.
Говоря о значении болезней различного происхождения в многолетней динамике численности лесных насекомых, следует отметить, что наибольшую роль играют так называемые смешанные заболевания, которые вызываются не одним, а несколькими возбудителями, например вирусом и простейшими, вирусом и энтомофторовыми грибами. В этом случае может наблюдаться сокращение сроков развития отдельных фаз вспышки массового размножения насекомого.
Паразитические насекомые как естественные враги других насекомых особенно многочисленны. К их числу относятся такие обширные группы, как наездники, хальцидовые из перепончатокрылых, тахины и серые мясные мухи из двукрылых, нарывники из жесткокрылых. По образу жизни и типу питания паразиты делятся на эктопаразитов (внешних) и эндопаразитов (внутренних). Первые живут на теле хозяина снаружи и питаются через его покровы; вторые развиваются внутри тела хозяина).
Существуют различные способы откладки яиц паразитами: откладка яйца внутрь тела насекомого, на поверхность кожных покровов хозяина, на кормовой субстрат хозяина – в этом случае яйцо проглатывается насекомым вместе с кормом.
В зависимости от того, на какой стадии жизненного цикла хозяина развивается паразит, различают: яйцеедов (весь цикл развития эти насекомые проходят в яйце хозяина); гусеничных паразитов (паразит заражает гусеницу хозяина, проходит цикл развития и выходит из гусеницы в более позднем ее возрасте); гусенично-куколочных паразитов (заражают хозяина на стадии гусеницы и выходят из куколки); куколочных паразитов (заражение и выход паразита проходит на стадии куколки хозяина).
Насекомое - хозяин является для паразита не только источником питания, но и средой обитания. Вследствие этого между их жизненными циклами устанавливается согласованность. Это достигается путем синхронизации сроков появления имаго паразита и заражаемой фазы развития хозяина, одинакового числа генераций и совпадения сроков наступления диапаузы.
По степени специализации энтомофаги подразделяются на монофагов – приспособленных к питанию на одном виде насекомых; олигофагов - развивающихся на ограниченном количестве близких видов и полифагов – заражающих большое число видов насекомых.
В фазе имаго паразиты ведут свободный образ жизни и, обычно, нуждаются в дополнительном питании. С этой целью они используют пыльцу и нектар цветов, либо клеточный сок, выступающий из ранок растений, либо гемолимфу будущего хозяина, выступающую из специально нанесенных ранок.
Кроме паразитов первого порядка, развивающихся за счет растительноядных насекомых, существуют паразиты второго порядка, жизненный цикл которых проходит в различных фазах онтогенеза паразитов первого порядка. При этом вторичные паразиты, уничтожая первичных паразитов, снижают уровень влияния их на хозяина. Существуют также паразиты более высоких порядков – третьего, четвертого, пятого и даже шестого порядков. Всех паразитов, начиная с паразитов второго порядка, принято называть сверхпаразитами.
Возможно явление сопаразитизма, когда в одну особь хозяина откладываются яйца нескольких видов паразитов. При этом наблюдается эффект межвидовой конкуренции паразитов. В результате конкуренции за место и пищу один паразит может погибнуть, а другой выжить. Могут выжить оба паразита, однако, в этом случае оба они, как правило, характеризуются низкой плодовитостью и малыми размерами. Может наблюдаться и гибель обоих паразитов вследствие недостатка корма или ранней гибели хозяина.
Следует отметить, что насекомые обладают рядом защитных приспособлений против паразитов. К числу механических реакций можно отнести резкие движения тела или конечностей насекомого, отпугивающие паразитов, бегство от паразита, стряхивание паразита с поверхности тела и т.д.
Из физиологических защитных реакций наиболее изученными являются инкапсуляция яиц и личинок клетками гемолимфы хозяина. Полное образование капсулы завершается за несколько дней и паразит гибнет. Физиологическая защита может активно проявляться лишь на начальных этапах подъема численности насекомого, когда особи еще не ослаблены недостатком корма. Против специализированных паразитов у многих насекомых гемоцитарная реакция вообще отсутствует.
В большинстве случаев жизненный цикл лесных насекомых длится один год. Примером может служить такой опасный вредителей лесов, как непарный шелкопряд Lymantria dispar L. (сем. Lymantriidae, отр. Lepidoptera). Весной из отложенных осенью прошлого года яиц выходят гусеницы, которые, пройдя ряд последовательных линек, окукливаются. Через некоторое время из куколок вылетают бабочки, которые спариваются, и затем самки откладывают яйца. Таким образом, развитие стадий гусеницы, куколки и имаго у этого вида длится с мая по август одного календарного года.
Более сложным является цикл сезонного развития сибирского шелкопряда Dendrolimus superans sibiricus Tschetv. (сем. Lasiocampidae, отр. Lepidoptera), у которого, в зависимости от фазы градации, цикл развития особей может длиться один или два года.
В течение всего периода сезонного развития насекомых численность их популяций может значительно изменяться. В каждый момент времени состояние популяции можно выразить в виде набора цифр, характеризующих численность (плотность) особей, находящихся на определенной стадии жизненного развития (яйца, личинки, куколки, имаго), который оформляется в виде таблиц, называемых таблицами выживаемости. Таблицы выживаемости отражают интенсивность влияния различных экологических факторов на изменение численности популяции на определенной стадии сезонного развития.
Способы построения таблиц выживаемости
В целях оценки выживаемости особей определенного вида на разных фазах развития выборка из определенного числа особей изолируется в природных или лабораторных условиях. Дальнейшие наблюдения ведутся за одними и теми же особями. Для этих особей фиксируются моменты гибели и моменты перехода в следующую стадию развития, определяются факторы смертности. На основании подобных наблюдений и составляются таблицы выживания. В настоящее время таблицы выживаемости построены для многих насекомых–вредителей леса. В качестве примера приводится таблица выживаемости (таблица 6.2) для бабочки Choristoneura fumiferana (сем.Tortricidae), живущей в лиственничных насаждениях Канады.
В таблице 6.2 показатель lx соответствует численности популяции в начале каждой стадии (здесь, например, в кладке 200 яиц), dx - смертности в течение времени x.
Таблица 6.2 - Таблица выживаемости для Choristoneura fumiferana
| Стадии развития | lx | Фактор смертности | dx | dx % от lx |
| Яйца | Паразиты | |||
| Прочие | ||||
| Всего | ||||
| Молодые личинки | Рассеяние | |||
| Более взрослые личинки | Паразиты | 13,6 | ||
| Болезни | 6,8 | |||
| Прочие | 10,2 | |||
| Всего | 30,6 | |||
| Куколки | 3,4 | Паразиты | 0,35 | |
| Прочие | 0,55 | |||
| Всего | 0,9 | |||
| Имаго | 2,5 | Разные | 0,5 | |
| Соотношение полов 1:1 | Смертность 99%, | Выжива-ние 1% |
Из таблицы видно, что из 200 яиц выходит всего 2 особи взрослых насекомых, т.е. смертность равна 99%. Наиболее уязвимой стадией являются молодые личинки, так как их смертность достигает 80%. Если принять, что соотношение полов у данного вида равно единице, то при сохранении постоянной численности популяции смертность должна составлять:
- при 20 потомках на одну самку – 90%;
- при 200 потомках на одну самку – 99%;
- при 2000 потомках на одну самку – 99,9%.
Особенностью таблиц выживаемости, подобных приведенной выше, является неявное предположение о том, что ее коэффициенты не зависят от времени и не меняются от сезона к сезону. Если это предположение неверно, то для каждого сезона нужно строить свою таблицу выживаемости, что крайне усложняет прогнозирование численности популяции. К сожалению, для видов, дающих вспышки массового размножения (то есть для наиболее интересных с точки зрения специалистов-энтомологов), предположение о постоянстве коэффициентов таблицы выживаемости неверно (так, на фазе кризиса доля особей в популяции, погибших вследствие воздействия паразитов и хищников, резко возрастает).
Наиболее сложно строить таблицу выживаемости, когда нет никакой возможности выращивать особей из популяции в лабораторных условиях. Так, это невозможно при построении таблицы выживаемости насекомых-минеров.
У большинства видов лесных насекомых сезонный цикл развития длится не меньше года. Поэтому для оценки особенностей многолетней динамики численности популяций необходимы длительные наблюдения за этими популяциями.
Самый «длинный» ряд многолетней динамики численности включает в себя данные по динамике серой лиственничной листовертки Zeiraphera griseana Hbn. (сем. Tortricidae, отр. Lepidoptera) в Швейцарских Альпах за период более чем в 150 лет. Сопоставимы по своему объему с данными по серой лиственничной листовертке данные по динамике численности трех видов насекомых – филлофагов в Германии (область Тюрингия): сосновой пяденицы (Bupalus piniarius L., сем. Geometridae, отр. Lepidoptera.), соснового шелкопряда (Dendrolimus pini L., сем. Lasiocampidae, отр. Lepidoptera.), сосновой совки (Panolis flammea Den. et Schiff., сем. Noctuidae, отр. Lepidoptera). Непрерывные наблюдения за популяциями этих видов велись в течение 90 с лишнем лет.
Форма представления данных по динамике численности популяции может быть различной. Например, данные можно представить в табличной форме, где в одной из колонок таблицы указывается год учета, а в другой - численность (плотность) популяции данного вида. Однако табличная форма представления данных о динамике численности популяций не очень удобна. Длинные ряды представлять в таблице неудобно технически (трудно воспринимать таблицу со 150 строками!). Кроме того, для специалиста наибольший интерес представляют не абсолютные значения плотности популяции, а динамика изменения плотности популяции во времени. Но в табличной форме представления данных такой анализ затруднителен.
Более удобно анализировать данные по динамике численности, представленные в графической форме. Рассмотрим для примера данные по динамике численности опасного вредителя лесов Северной Америки – еловой листовертки (Choristoneura fumiferana Clem., Lepidoptera, Tortricidae). В таблице 6.3 приведены данные учетов плотности популяции этого вида на различных фазах градации (Уатт, 1971).
Таблица 6.3 -Учеты плотности популяции еловой листовертки в Канаде
| Год | ||||||
| плотность популяции, гусениц/дерево | 3,1 | |||||
| Год | ||||||
| плотность популяции, гусениц/дерево | 3,3 |
Существуют три способа графического представления данных о динамике численности популяций.
Представление в плоскости (численность-время). В этом случае по оси Х (в этом случае это ось времени t) откладываются годы (если мы имеем дело с популяцией, учеты численности которой проводятся один раз в год), а по оси Y откладываются величины плотности популяции в годы учетов (рисунок 6.2).
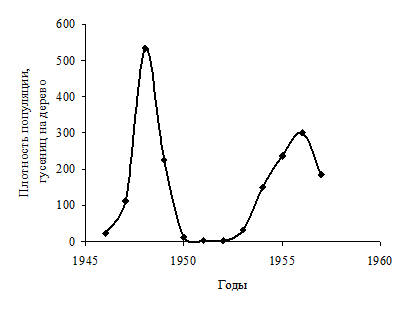
Рисунок 6.2. - Временная динамика численности популяции еловой листовертки
Представление в плоскости (численность i-го поколения - численность (i+1)-го поколения). В этом случае по оси Х откладываются значения плотности популяции в i-ом году, а по оси Y – величины плотности популяции в (i+1)-ом году. Способ построения графика состоит в следующем: выбираем пару данных о численности популяции в первом и втором годах учетов. Таким образом, получаем первую точку графика, у которой данные для первого года есть значение по оси Х, а данные для второго года – значение по оси Y. Затем, берем пару (данные второго года, данные третьего года) и для этой пары строим точку на плоскости, аналогично паре данным для первого и второго годов. Точку 1 соединяем с точкой 2. Затем, эту процедуру повторяем для всех смежных пар данных. В результате получим набор точек, соединенных друг с другом линиями. Получившаяся ломаная линия характеризует изменения динамики численности исследуемого вида во времени. Такой способ представления носит название лестницы Ламерея (рисунок 6.3).
Предположим, что численность популяции остается постоянной в течение всего времени учетов. Тогда все величины численности (плотности популяции) одинаковы для всех моментов времени t. Значения X и Y для всех точек строящегося графика также будут равными, а отношение Y к X всегда будут равны единице. Таким образом, в этом случае в плоскости (численность i-го поколения - численность (i+1)-го поколения) многолетняя динамика численности популяции будет характеризоваться всего одной точкой, расположенной на биссектрисе Y = X.
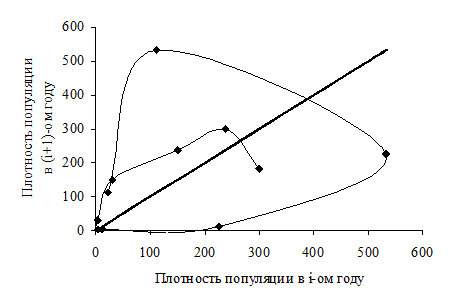
Рисунок 6.3 - Представление популяционной динамики еловой листовертки в форме лестницы Ламерея
Представление в плоскости (численность – коэффициент размножения). При таком способе представления динамики численности вычисляется коэффициент размножения y(t) популяции с неперекрывающимися поколениями – отношение плотности популяции Y(t+1) в момент времени (t+1) к плотности популяции Y(t) в момент времени t:  .
.
Следует отметить, что для расчетов коэффициента размножения необходимы данные учетов численности насекомых двух смежных лет (поколений), поэтому по однократному учету численности популяции изучаемого вида насекомых рассчитать коэффициент размножения невозможно.
Для описания динамики численности популяции в плоскости (численность – коэффициент размножения) или, как ее еще называют, в фазовой плоскости, по оси Х откладываются значения численности популяции Y(t) в момент времени t, а по оси Y – величина коэффициента размножения y(t). Затем строится следующая точка, характеризующая численность популяции и коэффициент размножения в следующий момент времени. Две этих точки соединяются прямой. Если учеты численности проводились в течение n лет, то число точек на фазовой плоскости будет равно n – 1, так как для последнего значения плотности популяции невозможно вычислить коэффициент размножения. Кривая на фазовой плоскости, характеризующая связь между плотностью популяции и коэффициентом размножения, носит название фазовой траектории популяции или фазового портрета (рисунок 6.4).

Рисунок 6.4 - Фазовый портрет популяции еловой листовертки
Точно так же, как при представлении в плоскости (численность i-го поколения - численность (i+1)-го поколения), существует выделенное место точек – биссектриса, точки на которой характеризуют популяцию, численность которой не меняется в течение двух смежных моментов времени, на фазовой плоскости существует аналог биссектрисы – прямая y(t) º 1 или же, если для удобства представления рассматриваются не величины плотности популяции и коэффициенты размножения, а их логарифмы, прямая ln y(t) = 0. Если какая-либо точка попадает на эту прямую, это означает, что коэффициент размножения для двух смежных поколений равен единице и, следовательно, численности двух смежных поколений популяции насекомых равны.
В фазовой плоскости популяция со стабильным типом динамики численности характеризуется просто одной точкой, популяция со ступенчатым типом динамики численности – тремя точками.
Всегда возможно перевести показатели динамики численности популяции, представленные с помощью одного способа, в другой способ представления.
| <== предыдущая страница | | | следующая страница ==> |
| Качественные популяционные показатели | | | Типы многолетней динамики численности популяций лесных насекомых |
Дата добавления: 2014-02-27; просмотров: 1388; Нарушение авторских прав

Мы поможем в написании ваших работ!